
Antibodi atau imunoglobulin merupakan protein yang berperan penting dalam respon imunitas adaptif. Peranannya terutama mengenali antigen dan kemudian menjembatani proses pengenalan dengan netralisasi atau pembersihan antigen tersebut dari tubuh. Pada kesempatan kali ini kita akan membahas lebih detil mengenai struktur dan fungsi dari antibodi tersebut.
Daftar Isi
Struktur Molekul Antibodi
Antibodi merupakan protein dengan bentuk kasar seperti huruf “Y”. Bagian atas dari imunoglobulin yang berbentuk cabang merupakan regio yang bertanggung jawab untuk mengikat antigen dan dinamakan binding region (Fab). Bagian Fab ini tersusun atas subprotein rantai berat/heavy chain (HC) dan subunit protein rantai ringan/light chain (LC). Keduanya memiliki variabiltias struktur molekul dan menentukan spesivisitas maupun afintias dari antibodi.
Adapun bagian ekor dari molekul imunoglobulin dinamakan regio Fc. Regio ini bertugas beinteraksi dengan komponen sistem pertahanan tubuh seperti reseptor di permukaan sel dan komplemen. Dengan interaksi tersebut, imunoglobulin akan mengaktivasi proses sistem imunitas lanjutan seperti pelepasan sitokin, mendorong maturasi sel, fagositosis antigen, dan sebagainya. Di bawah ini adalah gambar dari struktur molekul dan diagram dari struktur antobodi.


Struktur Fab
Bagian Fab yang memiliki variabilitas untuk mengenali antigen terdapat di regio variabel atau regio V. Regio ini penting dalam pengenalan antigen sehingga menjadi fokus penelitian dalam menyusun antibodi monoklonal untuk tujuan terapi maupun mendesain antigen yang digunakan sebagai vaksin. Regio V ini terdiri dari enam bagian complementarity-determining region (CDR) yakni tiga dari subunit HC dan tiga dari subunit LC. Residu asam amino yang berkontak langsung dengan antigen disebut dengan istilah paratope. Adapun permukaan antigen yang berkontak langsung dengan antibodi dikenal sebagai epitope.
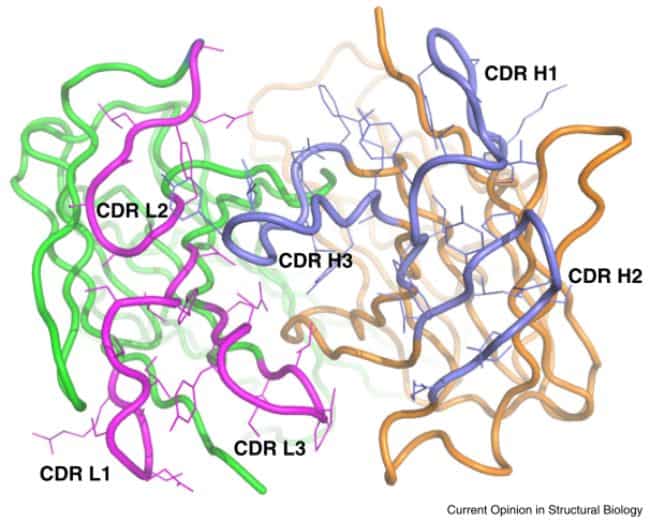
Struktur Fc
Adapun bagian bawah dari imunoglobulin, dinamakan bagian atau regio Fc. Fc ini terdiri dari sepasang subunit protein HC dengan struktur yang konstan atau tetap. Bagian Fc bertugas sebagai fungsi efektor dan berperan dalam menentukan waktu paruh dari antibodi. Fungsi efektor dari Fc termasuk terlibat dalam mekanisme pembunuhan sel melalui interaksi protein dan karbohidrat dengan reseptor Fcγ maupun sistem komplemen. Adapun fungsi penentuan waktu paruh berkaitan dengan interaksi Fc dengan FcRn.
Mengenai pembahasan struktur molekul antibodi atau imunoglobulin juga dapat disimak di video di bawah ini:
Reseptor Fc (FCR)
Seperti disebutkan di atas, bagian Fc dari imunoglobulin akan berinteraksi dengan efektor melalui reseptor Fc. Karena sebagian besar antibodi berupa IgG, kita akan melakukan penjelasan lebih rinci dari reseptor Fcγ (FcγR). FcγR meliputi beberapa jenis anggota reseptor dengan pola ekspresi berikut ini:

Dari bagan di atas tampak bahwa setiap sel memiliki jenis dan komposisi kelas FcγR yang berbeda. Selain itu, FcγR ada yang bersifat menginduksi ekspresi sistem imun dan ada yang merepesi atau mengurangi respon dari sistem imun.
Jalur Sinyal FcγR
Walaupun terdapat perbedaan struktur antara anggota keluarga FcγR, namun semuanya memiliki pola sinyal transduksi yang sama saat reseptor diaktivasi. Kecuali FcγRI, semua anggota dari FcγR lainnya memiliki afinitas yang rendah terhadap IgG dan hanya dapat berinteraksi dengan kompleks IgG multimer atau saat antigen teropsonisasi pada waktu peristiwa infeksi. Adapun FcγRI bisa berinteraksi dengan IgG monomer dengan afinitas yang tinggi. Oleh sebab itu, walaupun terdapat kadar antibodi tinggi di dalam serum, namun tanpa keberadaan pemicu patogenik, antibodi tidak mampu berikatan dengan FcγR sehingga menghindari aktivasi sel efektor yang tidak perlu.
Adapun apabila terdapat antigen, maka kompleks antigen-antibodi dapat mengadakan crosslinking dengan FcγR sehingga dapat mengaktivasi efektor. Crosslinking ini menyebabkan reseptor berkumpul atau beragregarsi dan kemudian akan memfosforilasi domain ITAM – motif tandem Yxx/L. Fosforilasi ini dilakukan oleh keluarga kinase SRC (misalnya LYN, LCK, HCK, dan FGR) maupun melalui aktivasi dari keluarga kinase SYK.
Langkah penting dari peristiwa kaskade fosforilasi ini adalah aktivasi dari PI3K oleh SYK yang kemudian akan merekruit pleckstrin homology domain-expressing protein seperti BTK, GAB2, dan phosphoinositide-specific phospholipase Cγ (PLCγ). Protein-protein ini membantu pembentukan inositol triphosphate (IP3) yang kemudian akan memobilisasi ion kalsium (Ca2+) dari retikulum endoplasma dan diacylglycerol (DAG). Baik Ca2+ dan DAG kemudian akan mengaktivasi protein kinase C (PKC). Untuk fungsi fosforilasi dapat dibaca di artikel ini.
PAC kemudian akan menyebabkan rangkaian aktivasi Rho GTPases CDC2, RAC1, RAC2, dan polimerisasi aktin oleh protein ARP2/3 dan WASP. Hasilnya adalah fagositosis dari kompleks IgG dan internalisasi dari reseptor. Selain dari jalur persinyalan ini, secara bersamaan juga terjadi aktivasi jalur lain termasuk MEK dan keluarga MAP kinase dan RAS. Proses ini akan menyebabkan ekspresi sitokin proinflamasi dan berbagai kemokin.
Kejadian di atas akan diimbangi oleh aktivitas dari FcγRIIb yang dimediasi oleh rekrutmen fosfatase pada domain ITIM diikuti crosslinking dari reseptor serta fosforilasi oleh keluarga kinase SRC. Fosfatase yang direkrut ITIM seperti SHP1 dan SHP2 akan mempromosikan hidrolisis phosphatidylinositol 3,4,5- triphosphate (PIP3) menjadi phosphatidyinositol 4,5- biphosphate (PIP2). Peristiwa hidrolisis ini terjadi di bagian sisi dalam dari membran sel dan efeknya adalah menghambat rekrutmen serta aktivasi dari PLCγ serta tirosin kinase BTK. Dikarenakan pada leukosit memiliki baik FCγR maupun FcγRIIb, maka hasil akhir dari aktivasi proinflamasi atau kontrainflamasi tergantung dari keseimbangan antara kedua jenis reseptor induser maupun inhibitor yang diaktivasi di permukaan sel.

Fungsi Efektor yang Dimediasi Antibodi
Saat terjadi aktivasi efektor, terjadi beberpa fungsi yang berhubungan dengan aktivitas pertahanan tubuh. Adapun fungsi-fungsi tersebut diantaranya adalah sebagai berikut:
Degranulasi dan Respiratory Burst
Efek aktivasi dari FcγR bisa berlainan dari sel ke sel. Pada granulosit seperti neutrofil, basofil, dan eosinofil, aktivasi dari kinase SYK dan SCR akan menyebabkan penyusunan kompleks NADPH-dependent oxidase di membran plasma dan membran dari fagosom. Hal ini akan memproduksi reactive oxygen species dan reactive nitrogen species yang keduanya memiliki kemampuan untuk membunuh mikroorganisme. Selain itu, aktivasi PKC dan peningkatan kadar Ca2+ intraseluler memicu mobilisasi dan pelepasan secara cepat isi granul dari sel granulosit. Isi granul ini berupa protease serine, leukotrien, lisozim, lactoferin, dan α-defensin. Isi dari granul tersebut memiliki kemampuan dalam membunuh mikroorganisme.
Bersihan/Clearance Virion yang Teropsonisasi
Selain degranulasi, aktivasi FcγR pada neutrofil dan makrofag juga akan memicu fagositosis antigen yang teropsonisasi (antigen yang dikelilingi antobodi). Gambar di bawah ini menggambarkan bagaimana neutrofil maupun makrofag berepson terhadap virion yang teropsonisasi.

Eliminasi Sitotoksik dan Fagositosis dari Sel yang Terinfeksi Virus
Sebagai mekanisme pertahanan tubuh terhadap infeksi virus, sel yang terinfeksi virus sering kali mengekspresikan antigen virus di permukaan selnya. Antigen ini dapat dikenali oleh antibodi dan kemudian bagian Fc dari antibodi dapat berinteraksi dengan FcγR dari sel NK. Hal ini akan menyebabkan sel NK menjadi aktif dan melakukan degranulasi untuk membunuh sel yang terinfeksi virus.

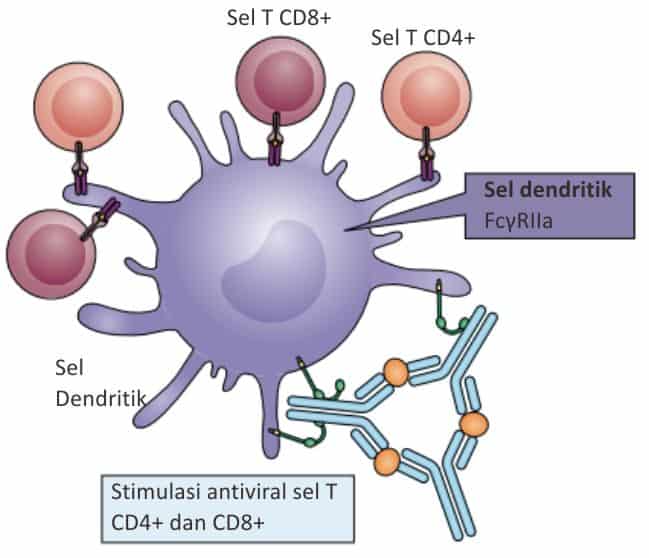
Efisiensi Fungsi Antigen Presenting Cell (APC)
Crosslinking dari FcγR pada sel dendritik akan mempercepat proses fusi dan maturasi fagolisosom. Hal ini akan menyebabkan pemrosesan dan presentasi antigen yang lebih cepat dan efisien.
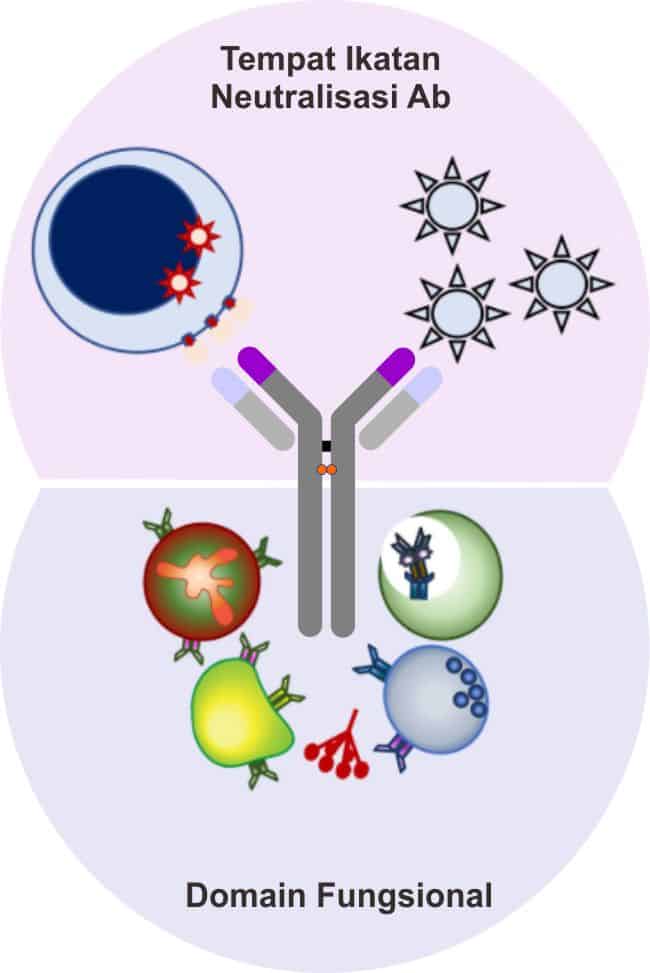
Glikosilasi dari Antibodi
Dari bagan diagram di atas, tampak bahwa pada molekul antibodi terdapat struktur karbohidrat. Penambahan struktur karbohidrat ini dinamakan proses glikosilasi. Adapun proses glikosilasi ini dapat terjadi di regio Fab, Fc, maupun di hinge region. Molekul karbohidrat yang menempel pada antibodi umumnya adalah oligosakarida berukuran besar (~2 kDa per oligosakarida), bersifat fleksibel, dan umumnya berikatan dengan antibodi melalui ikatan N-glikosidik maupun O-glikosidik.

Adapun fungsi dari struktur oligosakarida ini terutama berperan dalam proses ikatan antibodi dengan molekul lain seperti proses ikatan dengan leptin serum (MBL) serta berperan dalam menjaga kelarutan dan konformitas dari struktur antibodi. Fungsi lainnya meliputi fasilitasi transport, sekresi, serta cleareance subseluler; dan menjaga fungsi efektor dengan mempertahankan ikatan Fc dengen reseptor Fc yang optimal.
Peran Antibodi dalam Menetralisir Infeksi Virus
Sebenarnya fungsi antibodi ini sangat kompleks. Pengetahuan mengenai mekanisme antobodi memproteksi tubuh terhadap serangan berbagai macam infeksi telah berkembang dengan dengan sangat pesat. Akan tetapi, masih banyak area yang belum dipahami secara utuh. Berikut kita akan lihat apa saja yang telah kita ketahui mengenai antibodi baik dari studi in vitro (di cawan petri/di luar dari tubuh) maupun studi in vivo (fenomena fungsional di dalam sel/di dalam tubuh).

Aktivitas Antiviral Antibodi In Vitro
Seperti dijelaskan di atas, antibodi dapat memerangi virus yang beredar di dalam darah atau cairan tubuh maupun sel yang terinfeksi oleh virus. Barangkali peran yang sangat penting dari antibodi adalah peranannya dalam menetralisir partikel bebas dari virus.
Aktivitas Antiviral Antibodi Terhadap Partikel Bebas Virus
Partikel bebas virus berarti virus yang berada di luar sel. Adapun neutralisasi dari antibodi berarti hilangnya daya infeksi dari partikel virus saat molekul antibodi mengikat partikel virus, umumnya tanpa keterlibatan proses imunitas yang lain. Proses neutralisasi virus ini serupa dengan proses neutralisasi terhadap toksin dan enzim.
Adapun mekanisme pasti dari daya neutralisasi antibodi ini terhadap partikel bebas virus ternyata masih diperdebatkan. Hipotesis yang paling utama menyatakan bahwa ikatan antibodi akan merubah konformasi dari molekul permukaan virus. Adapun teori lain menyatakan bahwa antibodi berperan setelah virus masuk ke dalam sel dengan mencegah terjadinya uncoating dari envelop virus.
Porsi Fc dari imunoglobulin juga berperan dalam menetralisir partikel bebas virus. Beberapa meknismenya adalah sebagai berikut:
- Aktivasi komplemen oleh antibodi yang terikat kepada virus serta deposisi komplemen di permukaan virion. Hal ini akan mencegah pengikatan virus ke reseptor permukaan sel
- Aktivasi komplemen akan menyebabkan lisis virion secara langsung.
- Reseptor Fc dan komplemen dapat mengikat virion yang terikat antibodi dan/atau komplemen yang kemudian akan menyebabkan fagositosis yang diikuti oleh inaktivasi virion di dalam komponen intraseluler dari fagosit.
Terkadang pada virus tertentu di permukaan virion terdapat sedikit sekali antigen yang dapat diikat oleh antibodi. Pada kondisi tersebut, pengikatan antibodi tidak dapat menetralisir virus. Tetapi, antibodi tersebut masih dapat merusak virus melalui aktivasi sistem komplemen atau melalui proses fagositosis.
Aktivitas Antiviral Antibodi Terhadap Sel yang Terinfeksi Virus
Pada sel yang telah terinfeksi oleh virus, terikatnya antibodi pada permukaan sel dapat memicu beberapa aktivitas sistem imun (lihat gambar di atas). Sistem efektor yang diperantarai oleh Fc dapat menyebabkan lisis sel akibat antibody-dependent cellular cytotoxicity (ADCC) atau complement-dependent cytotoxicity (CDC).
Antibodi yang terikat molekul virus di permukaan sel yang terinfeksi juga dapat menghambat replikasi virus. Proses ini diduga disebabkan oleh mekanisme persinyalan sel. Fenomena ini terutama terobservasi pada infeksi sel saraf atau neuron. Selain itu, antibodi juga dapat menghambat pelepasan virion dari sel yang terinfeksi dan infeksi melalui perantara sel ke sel.
Akan tetapi, terdapat data bahwa fenomena antibodi terhadap sel yang terinfeksi lebih lemah dari pada terhadap partikel bebas virus. Misalnya diperlukan kadar antibodi yang lebih tinggi untuk menetralisir transmisi dari sel ke sel dibandingkan dengan untuk menetralisir partikel bebas virus.
Aktivitas Antiviral Antibodi In Vivo
Aktivitas in vivo artinya pengaruh sesuatu langsung di dalam tubuh. Studi in vivo antibodi sederhananya dengan memasukan antibodi ke hewan model baik berupa antibodi monoklonal maupun serum kemudian hewan coba terebut dipaparkan terhadap agen infeksius. Setelah itu dilihat reaksi yang terjadi termasuk apakah penyakit akan muncul pada hewan coba tersebut.
Dari data memang terjadi keseuaian antara aktivitas in vitro dengan in vivo. Antibodi yang bersifat menetralisir secara in vitro biasanya memberikan daya proteksi secara in vivo terhadap penyakit. Namun, tentu hal ini tidak 100% seperti itu dan metode netralisasi oleh antibodi bukan merupakan mekanisme satu-satunya dalam melindungi tubuh dari infeksi.
Biasanya, antibodi akan memberikan proteksi apabila kadar atau titernya sangat tinggi dalam darah saat terjadinya kejadian infeksi. Yang dimaksud kadar tinggi biasanya adalah titer 90% yaitu dalam orde 1 diantara 100. Artinya, walaupun serum didilusi atau diencerkan 100 kali tetap memberikan 90% netralisasi secara in vitro. Cara lain untuk menggambarkan efektivitas virus secara in vivo adalah istilah steril, artinya dengan titer tertentu tidak terobservasi adanya replikasi virus pada hewan coba tersebut setelah dipaparkan pada agen infeksius.
Dari studi in vivo inilah kita mengetahui bahwa mekanisme protektif antibodi pada infeksi virus tidak hanya dari sifat kemampuan netralisasi saja. Contohnya, saat antibodi dipotong dan hanya berupa fragmen Fab tanpa bagian Fc, secara in vitro antibodi ini dapat memperlihatkan efek netralisasi. Akan tetapi, apabila fragmen Fab ini duji coba secara in vivo, ternyata tidak memberi dampak proteksi bagi hewan coba terhadap infeksi.
Aktivitas Antiviral dari Sel T
Menguji coba dengan mentransferkan secara pasif antibodi ke hewan coba dapat memberikan informasi mengenai daya proteksi suatu jenis antibodi. Namun, hal tersebut tidak memberikan gambaran yang utuh. Hal ini dikarenakan dengan melakukan transfer pasif, tidak mengaktivasi respon imunitas seluler maupun respon sekunder dari sel B.
Selain dari antibodi, sel T penting dalam tubuh untuk melawan infeksi virus. Hal ini terutama melawan sel tubuh yang terinfeksi virus. Sel T CD8+ berperan dalam melawan infeksi virus melalui fungsi pengenalan spesifik terhadap antigen melalui T-cell receptor (TCR). Antigen berupa peptida virus biasanya akan diekspresikan di permukaan sel yang terinfeksi berikatan dengan molekul MHC kelas I.
Kompleks antigen dengan MHC kelas I inilah yang berinteraksi dengan TCR. Akibat interaksi ini sel T CD8+ kemudian akan membunuh sel yang terinfeksi virus tersebut baik dengan menginduksi apoptosis atau mengeluarkan sitokin antiviral untuk membersihkan virus. Sel T CD4+ juga menunjukan aktivitas antiviral secara langsung tetapi magnitud aktivitas antiviralnya tidak setinggi sel T CD8+ maupun sel B.
Untuk aktivitas spesifik sel T ini jelas tidak bisa diaktivasi semata oleh hanya pemberian antibodi secara pasif. Oleh sebab itu, vaksinasi bersifat melindungi tidak hanya karena dari perangsangan produksi antibodi semata namun juga mengaktivasi respon imun lain termasuk sel T.
Mekanisme Virus Mengelabui Daya Proteksi Antibodi
Banyak virus mampu mengelabui daya proteksi antibodi. Hal ini misalnya terlihat dari infeksi influenza yang dapat berlangsung berulang kali pada individu yang sama serta kegagalan uji vaksin HIV. Berikut adalah beberapa mekanisme cara virus mengelabui daya proteksi antibodi:
- Mutasi protein virion sehingga setiap kali terjadi mutasi antibodi yang sudah terbentuk tidak lagi bersifat protektif terhadap virus baru yang sudah mengalami mutasi.
- Memproduksi antigen yang dilepas ke plasma. Antigen yang dilepas ke plasma ini biasanya berupa glikoprotein permukaan sel/surface glycoprotein. Antigen lepasan ini berfungsi sebagai umpan. Antigen umpan ini akan menempel ke antibodi dan dapat mengurangi afinitas antibodi terhadap partikel bebas dari virus. Contoh virus yang memiliki kemampuan ini misalnya HIV-1 dan Ebola.
- Mengekspresikan molekul di permukaan virion yang meregulasi komplemen. Molekul permukaan regulator ini misalnya CD46 dapat mencegah virolisis yang disebabkan oleh aktivitas sistem komplemen.
- Memproduksi berbagai jenis protein yang tidak lengkap dan/atau misfolded. Protein ini juga bersifat umpan atau decoy dimana virus berusaha agar tubuh memproduksi antibodi yang tidak sesuai dengan struktur protein virus yang asli atau yang lengkap.
- Memproduksi antigen permukaan yang kaya akan glikoprotein, Struktur karbohidrat dari glikoprotein akan menyembunyikan bentuk protein sehingga mengurangi antigenisitas maupun imunogenisitas dari virus tersebut.
- Mengurangi jumlah antigen atau reseptor di permukaan virus sehingga hal ini juga akan mengurangi antigenisitas maupun imunogenisitas dari virus.
Jenis-jenis Antibodi pada Tubuh Manusia
Diketahui terdapat lima kelas antibodi manusia yaitu IgG, IgM, IgA, IgD, dan IgE. Perbedaan kelas ini berdasarkan jenis atau tipe rantai berat dari antibodi. Selain itu, pada IgG, IgA, dan IgD, perbatasan antara Fab dan Fc yaitu regio engsel (hinge region) memiliki struktur yang fleksibel sedangkan pada IgM dan IgE struktur regio engsel tersebut tersusun atas struktur yang lebih rigid. Profil singkat kelima kelas antibodi tersebut dapat disimak di bagan di bawah ini:

IgG
IgG merupakan jenis imunoglobulin terbanyak di dalam plasma dan meliputi 70-75% dari total antibodi di dalam tubuh. Ukuran IgG cukup besar dengan berat molekul sekitar 150 kDa. IgG tersusun atas empat rantai polipeptida dengan dua rantai berat tipe γ. IgG berperan aktif dalam proses imunitas dan berperan dalam proteksi jangka panjang. imunoglobulin IgG melindungi tubuh dari serangan bakteri, virus, menetralisir toksin bakteri, memicu aktivasi sistem komplemen, dan mengikat antigen untuk menambah efektivitas dari proses fagositosis. IgG dapat menembus sawar darah plasenta. Proses ini memungkinkan imunitas dari ibu dapat ditransfer ke bayi sambil menunggu proses maturitas dari imuntias dari bayi.
IgM
IgM juga bersirkulasi di dalam darah dan mewakili kira-kira 10% dari imunoglobulin total tubuh manusia. Antibodi ini memiliki rantai berat tipe μ. Di dalam plasma, IgM sering membentuk struktur pentamer dimana lima IgM berikatan bersama-sama. IgM disekresikan sel B saat pertama kali berespon terhadap inveksi bakteri atau virus. Walaupun memiliki afinitas yang lebih rendah dari IgG, namun IgM mempunyai afiditas yang lebih tinggi karena struktur pentamer tersebut. IgM beperan dalam pengenalan antigen golongan darah ABO pada permukaan eritrosit. Selain itu, IgM akan berikatan dengan permukaan sel dan mengaktivasi jalur persinyalan dari sel. IgM juga berperan dalam peningkatan proses fagositosis.
IgA
IgA tersusun atas rantai berat tipe α (alpha). IgA kaya di serum, mukus hidung, air liur, air susu ibu, dan cairan di saluran pencernaan. IgA mewakili 10-15% dari total antibodi dalam tubuh manusia. Biasanya IgA akan membentuk dimer. IgA dari ASI akan masuk ke saluran cerna bayi dan memproteksi saluran cerna bayi dari infeksi berbagai jenis patogen.
IgE
Jenis imunoglobulin selanjutnya adalah IgE. Struktur rantai berat dari IgE adalah tipe ε (epsilon). IgE beredar dengan kadar yang sedikit, tidak lebih dari 0,001% dari total antibodi tubuh. Sebagian IgE berada di permukaan sel, terutama mastosit dan basofil. Fungsi fisiologisnya adalah melindungi tubuh dari infeksi parasit. Akan tetapi, ditempat dengan insiden infeksi parasit rendah, IgG berperan terutama pada reaksi alergi.

IgD
Kelas terakhir dari imunoglobulin manusia adalah IgD. IgD tersusun dari rantai berat tipe δ (delta). IgD diekspresikan terutama di permukaan membran plasma dari sel limfosit B matur. Kadarnya kurang dari 1% dari jumlah total antibodi tubuh manusia. Fungsi utamanya tidak diketahui namun dipercayai berperan dalam induksi produksi antibodi oleh sel limfosit B.
Perpindahan Kelas Antibodi (Immunoglobulin Class Switching)
Terdapat fenomena dimana sel B dapat berpindah dari awalnya memproduksi IgM dan IgD kemudian memproduksi antibodi kelas lainnya. Peristiwa ini dikenal sebagai immunoglobulin class switching. Biasanya pada awalnya terdapat sel B yang mengekspresikan IgG dan IgD di permukaan selnya. Kemudian, setelah terpapar oleh antigen spesifik, sel B akan berproliferasi dan mesekresikan IgM dan IgD ke darah. Paparan antigen spesifik lebih lanjut akan menyebabkan diferensi lebih lanjut dan sel B tersebut akan meningkatkan produksi imunoglobulin dan mulai memproduksi kelas imunoglobulin lain selain dari IgM dan IgD. Contohnya adalah sebagai berikut:
- Interleukin-4 (IL-4) menstimulasi pergantian kelas dari IgM/IgD menjadi IgG1 dan IgE
- IL-5 menstimulasi pergantian kelas dari IgM/igD menjadi IgA

Antibody-Dependent Enhancement (ADE)
ADE merupakan suatu fenomena dimana patogenesis dari sebagian infeksi virus akan meningkat dengan adanya antibodi sub-neutralizing atau antibodi non-neutralizing yang bereaksi silang. Dalam kata lain, dalam kondisi tertentu bisa diproduksi antibodi yang sifat neutralisasinya kurang kuat atau terdapat reaksi silang antibodi dari infeksi virus jenis lain. Namun, alih-alih antibodi ini melawan infeksi virus, antibodi sub-neutralizing atau non-neutralizing ini malah membantu virus menginfeksi tubuh dengan lebih cepat.
Contoh yang sering kita hadapi adalah fenomena ADE pada infeksi virus Dengue atau penyakit demam berdarah. Kita ketahui terdapat empat serotipe dari virus Dengue yaitu DENV-1, DENV-2, DENV-3, dan DENV-4. Apabila seseorang terinfeksi salah satu serotipe misalkan DENV-1, maka tubuh orang tersebut akan memproduksi antibodi terhadap DENV-1. Karena ada kemiripan dengan serotipe lainnya, maka antibodi terhadap DENV-1 juga akan bereaksi silang terhadap virus serotipe lainnya.
Namun, antibodi yang bereaksi silang ini terhadap serotipe lainnya hanya bersifat menetralisi sebagian (sub-neutralizing atau non-neutralizing). Efeknya adalah timbulnya fenomena ADE. Pada kenyataannya, memang apabila orang yang pernah terinfeksi DENV-1 kemudian terinfeksi kedua kalinya dengan virus dengue serotipe lain, umumnya pada infeksi kedua gejala dan klinis penyakitnya akan lebih parah.
Adapun mekanisme penyebabnya pada infeksi dengue adalah saat virus terikat oleh antibodi, maka kompleks virus dengan antibodi ini akan berinteraksi dengan FcγR. Interaksi ini akan menyebabkan internalisasi atau virus dibawa masuk ke sel retikuloendotel (makrofag, endotel, neutrodil, dll). Karena sifatnya yang hanya menetralisir sebagian, proses internalisasi ini justru akan mempercepat proses infeksi virus sehingga klinis atau gejala yang timbul akan lebih berat dari sebelumnya.
Selain Dengue, beberapa virus lain juga menunjukan efek ADE ini. Diantaranya seperti influenza, HIV, dan Ebola. Untuk lebih lanjut mengenai ADE ini dapat disimak melalui artikel yang dikemukakan oleh Taylor A, et al.
Kesimpulan
Tidak diragukan lagi bahwa antibodi atau imunoglobulin sangat berperan sebagai lini pertahanan tubuh dalam melawan invasi dari berbagai jenis patogen. Perkembangan pengetahuan mengenai mekanisme proteksi antibodi ini sudah sangat berkembang. Akan tetapi, masih cukup banyak misteri yang belum terpecahkan dan membutuhkan penelitian yang lebih lanjut.
Referensi
- Arnold JN, Wormald MR, Sim RB, Rudd PM, Dwek RA. The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu Rev Immunol. 2007;25:21–50.
- Bournazos S, Gupta A, Ravetch J V. The role of IgG Fc receptors in antibody-dependent enhancement. Nat Rev Immunol. 2020;20(10):633–43.
- Burton DR. Antibodies, viruses and vaccines. Nat Rev Immunol. 2002;2(9):706–13.
- Chiu ML, Goulet DR, Teplyakov A, Gilliland GL. Antibody Structure and Function: The Basis for Engineering Therapeutics. Antibodies. 2019;8(4):55.
- Chiu ML, Gilliland GL. Engineering antibody therapeutics. Curr Opin Struct Biol. 2016 Jun;38:163–73.
- Cohen S. Antibody Structure. In: Antibodies for Infectious Diseases. American Society of Microbiology; 2015. p. 49–62.
- Taylor A, Foo SS, Bruzzone R, Vu Dinh L, King NJC, Mahalingam S. Fc receptors in antibody-dependent enhancement of viral infections. Immunol Rev. 2015;268(1):340–64.

Seorang dokter, saat ini sedang menjalani pendidikan dokter spesialis penyakit dalam FKUI. Peminat berbagai topik sejarah dan astronomi.
Comments 4
artikelnya mantap dan sangat jelas
Dokter Cecep Suryani Sobur, terima kasih banyak atas bantuannya.
Assalamu’alaikum Dokter Cecep Suryani Sobur, apakah memungkinkan untuk mendapatkan tulisan dokter dalam bentuk pdf. saya sangat senang membaca tulisan-tulisan dokter.
Author
Wlkmslm. Saya tidak menyimpan artikel dalam bentuk pdf karena saya langsung menulis dan mengedit di wordpress. Setahu saya bentuk artikel di browser bisa disimpan dalam bentuk pdf. Jadi bisa dicoba dibuka artikel terus langsung disimpan dalam bentuk pdf lewat broser chrome, firefox, dll