
Ekspresi gen merupakan proses sangat penting bagi tubuh. Proses ini dimulai dengan pembacaan gen melalui proses transkripsi DNA yang disalin menjadi RNA, dilanjutkan ke translasi RNA menjadi protein, dan kemudian dilakukan modifikasi pasca translasi atau modifikasi postranslasional. Pada artikel kali ini akan dibahas langkah awal dari ekspresi gen yaitu transkripsi DNA yang berupa sintesis RNA dan dilanjutkan dengan modifikasi RNA yang dihasilkan. Thumbnail oleh Gerd Altmann dari Pixabay

Selain di artikel ini, penjelasan mengenai transkripsi DNA juga dapat dibahas di video berikut ini:
Daftar Isi
Transkripsi DNA: Dari DNA Menjadi RNA
Pada transkripsi, DNA disalin ulang menjadi RNA. Pada dasarnya, kedua jenis molekul ini serupa. Baik DNA maupun RNA keduanya adalah polimer asam nukleat dengan tulang punggung gula fosfat dan tiap satu unit monomer memiliki gugus basa.
Perbedaan DNA dari RNA
Terdapat dua perbedaan penting antara DNA dan RNA. Perbedaan pertama terletak pada jenis gula dan perbedaan kedua adalah jenis gugus basa pirimidin timidin pada DNA menjadi urasil pada RNA.
Sesuai dengan namanya, gugus gula pada DNA adalah deoksiribosa sedangkan RNA gugus gulanya adalah ribosa. Deoksiribosa merupakan gugus gula ribosa yang kehilangan satu gugus hidroksil (OH). Perhatikan gambar di bawah ini:

Selain perbedaan di atas, pada dasarnya DNA dan RNA adalah sama. Begitu juga dengan pasangan basa urasil sama dengan thymine yaitu berpasangan dengan adenine.

Perbedaan Transkripsi DNA dengan Replikasi DNA
Serupa dengan proses replikasi DNA, pada transkripsi, RNA yang disintesis dibentuk berdasarkan template dari DNA induk. Namun, terdapat perbedaan-perbedaan yang mendasar.
Pada transkripsi, RNA yang terbentuk tidak akan tetap terikat dalam ikatan hidrogen dengan strand DNA template-nya. Tepat di belakang tempat ribonukleotida baru ditambahkan, rantai RNA akan terlepas dan bentuk heliks DNA akan segera terbentuk kembali. Oleh sebab itu, RNA yang terbentuk dilepas dari template sebagai single strand RNA.

Selain itu, tidak seluruh untai DNA ditranskripsikan namun hanya sebagian kecil saja. RNA yang terbentuk nantinya akan jauh lebih pendek. Sebagai contoh, satu molekul DNA pada kromosom manusia dapat mencapai 250 juta pasang basa nukleotida. Pada RNA panjangnya hanya beberapa ribu pasang nukleotida dan bahkan ada banyak yang lebih pendek dari itu.
Sifat lain dari RNA adalah kemampuan molekul ini untuk mengambil bentuk sekunder dan primer seperti protein. Dengan sifat ini, RNA dapat mengambil bentuk yang spesifik dan dapat juga membentuk situs katalitik sehingga RNA mampu berfungsi sebagai enzim layaknya protein. Perhatikan gambar di bawah ini mengenai fleksibilitas RNA dalam membentuk struktur spesifik:

RNA Polimerase: Enzim Utama pada Proses Sintesis RNA
Proses transkripsi DNA dijalankan oleh enzim RNA polimerase. Sama halnya dengan DNA polimerase pada proses replikasi DNA, RNA polimerase juga menjalankan reaksi ikatan fosfodiester yang menggabungkan satu nukleotida dengan nukleotida lainnya membentuk molekul polimer RNA. Gambar di bawah ini adalah posisi ikatan fosfodiester yang dibentuk dengan bantuan RNA polimerase:

Kemampuan enzimatik dari RNA polimerase dikarenakan memiliki bentuk unik berupa situs katalitik tempat terjadinya reaksi polimerisasi. Di tempat ini, molekul yang direaksikan ditempatkan saling berdekatan dan dengan bantuan atom magnesium reaksi pembentukan ikatan fosfodiester kemudian dapat terjadi. Untuk lebih memahami, perhatikan gambar di bawah mengenai struktur situs katalitik RNA polimerase:

Pada proses transkripsi, RNA polimerase berjalan sepanjang DNA dengan proses pelepasan struktur DNA heliks di depan situs akitf/situs katalitik. Adapun arah transkripsi adalah dari 5′ ke 3′. Adapun substratnya adalah nukleosida trifosfat yaitu ATP, CTP, UTP, dan GTP.
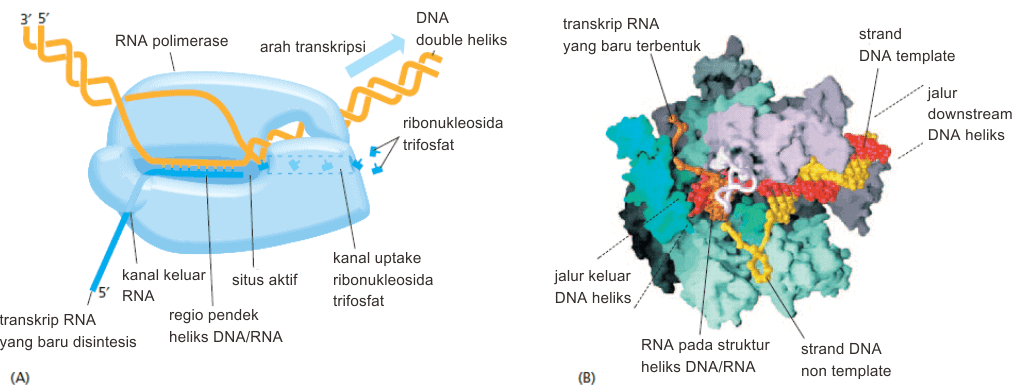
Dikarenakan strand RNA langsung dilepaskan dari template DNA-nya, maka hal ini memungkinkan beberapa salinan RNA dibuat secara bersamaan dari satu gen. Setiap RNA polimerase pada eukariota mampu menjalankan salinan 20 nukleotida per detik. Dengan cara ini, dari satu gen dapat dibuat ribuan transkrip dalam satu jam.

Perbedaan Reaksi RNA Polimerase dengan DNA Polimerase
Berikut adalah perbedaan-perbedaan reaksi yang dijalankan oleh RNA polimerase dengan DNA polimerase. Petama jelas, RNA polimerase mengkatalisasi pembentukan ikatan ribunukleotida, bukan deoksiribonukleotida.
Kedua, RNA polimerase dapat memulai proses transkripsi DNA tanpa memerlukan primer. Kemampuan ini diperkirakan dihubungkan dengan proses transkripsi DNA yang kurang memerlukan proses seakurat replikasi DNA.
Tidak seperti DNA, RNA tidak dipakai secara permanen untuk menyimpan informasi genetik (kecuali virus RNA). RNA polimerase membuat satu kesalahan untuk tiap 104 nukleotida yang disalin menjadi RNA (bandingkan DNA polimerase dengan satu kesalahan tiap 107 nukleotida yang disalin). Lagi pula, kesalahan transkripsi DNA tidak seberat kesalahan yang terjadi pada replikasi DNA.
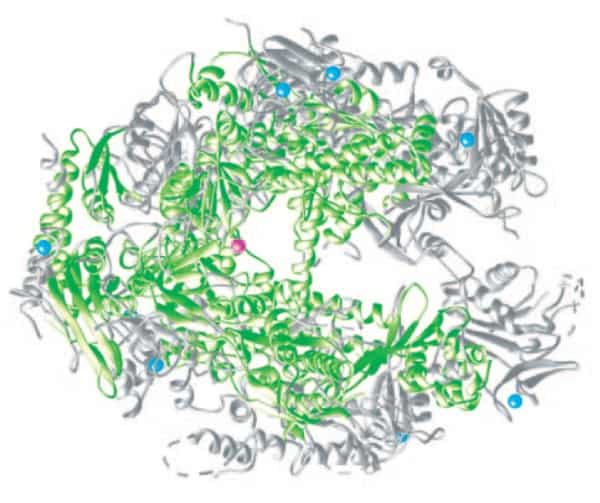
Secara struktur juga ternyata sangat berbeda. Walaupun sama-sama mengandung ion Mg2+ di situs katalitiknya, dari pembandingan kristalografi, bentuk kedua enzim itu tidak terkait satu sama lainnya. Di bawah ini adalah skema struktur RNA polimerase.

Adapun bentuk dasar dari DNA polimerase dapat dilihat di gambar di bawah ini:


Perbandingan Struktur RNA Polimerase Bakteri, Archaea, dan Eukariota
Kita tahu dalam taksonomi, makhluk hidup terbagi menjadi prokariota, archaea, dan eukariota. Membandingkan struktur RNA polimerase ketiga kingdom domain makhluk hidup ini sedikitnya memberi petunjuk proses evolusi makhluk hidup. Apabila kita perhatikan, struktur inti RNA polimerase terpelihara diketiga kingdom makhluk hidup ini. Perbedaannya adalah tambahan subunit dari enzim seperti tampak pada gambar perbandingan di bawah ini:

| Prokariota | Archaea | Euakriota RNAP I | Euakriota RNAP II | Euakriota RNAP III | |
|---|---|---|---|---|---|
| Subunit inti | β | Rpo1 | A190 | RPB1 | C160 |
| β’ | Rpo2 | A135 | RPB2 | C128 | |
| α | Rpo3 | AC40 | RPB3 | AC40 | |
| α | Rpo11 | AC19 | RPB11 | AC19 | |
| ω | Rpo6 | RPB6 | RPB6 | RPB6 | |
| Subunit spesifik archaea-eukariotik | Rpo5 | RPB5 | RPB5 | RPB5 | |
| Rpo8 | RPB8 | RPB8 | RPB8 | ||
| Rpo10 | RPB10 | RPB10 | RPB10 | ||
| Rpo12 | RPB12 | RPB12 | RPB12 | ||
| Rpo4 | A14 | RPB4 | C17 | ||
| Rpo7 | A43 | RPB7 | C25 | ||
| Subunit spesifik tipe RNAP | Rpo13 | ||||
| A12 | RPB9 | C11 | |||
| A49 | C53 | ||||
| A34.5 | C37 | ||||
| C82 | |||||
| C34 | |||||
| C31 |
Tampak bahwa subunit utama, yang dipertahankan mulai dari bakteri, archaea, maupun eukariota memiliki empat subunit. Subunit α2 yang merupakan dimer dari dua protein α, subunit β, subunit β’, dan subunit ω. Cara susunan empat subunit ini dapat disimak di gambar berikut:

RNA Polimerase pada Eukariota
Bakteri hanya memiliki satu tipe RNA polimerase. Pada eukariota, terdapat tiga jenis yaitu RNA polimerase I, RNA polimerase II, dan RNA polimerase III. Struktur ketiganya memiliki kemiripan namun mentranskripsikan tipe gen yang berbeda. RNA polimerase I dan III mentranskripsikan gen yang mengkode transfer RNA, ribosomal RNA, dan berbagai jenis small RNA. RNA polimerase II mengtranskripsikan sebagian besar gen termasuk seluruh gen yang mengkodekan protein.
| Tipe polimerase | Gen yang ditranskripsikan |
|---|---|
| RNA polimerase I | Gen rRNA 5.8S, 18S, dan 28S |
| RNA polimerase II | Semua gen yang mengkodekan protein, ditambah gen snoRNA, miRNA, siRNA, dan kebanyakan gen snRNA |
| RNA polimerase III | gen tRNA, 55 rRNA, beberapa snRNA, dan gen untuk small RNA lainnya |
Walaupun antara RNA polimerase II eukariota memiliki banyak kesamaan struktur dengan RNA polimerase bakteri, terdapat dua perbedaan yang mencolok:
- RNA polimerase hanya memerlukan tambahan satu protein (faktor σ) untuk inisiasi transkripsi, sedangkan RNA polimerase II memerlukan banyak tambahan protein yang dinamakan faktor transkripsi umum (general transcription factors)
- Inisiasi transkripsi pada eukariota harus berhadapan dengan struktur penyusun nukleosom DNA dan struktur kromatin lainnya. Hal ini tidak dijumpai pada kromosom bakteri.
Tipe atau Jenis RNA yang Diproduksi Sel
Sel ternyata tidak hanya memproduksi satu jenis RNA. Sebagian besar gen ditranskripsikan menjadi RNA dan kemudian menjadi template untuk sintesis protein. RNA yang semacam ini disebut messenger RNA (mRNA).
Sebagian kecil dari gen hanya menghasilkan RNA saja tanpa dilanjutkan seterusnya ke sintesis protein. Misalnya pada ragi, S. cerevisiae memiliki kurang lebih 750 gen (10% dari gen total) yang menghasilkan RNA saja sebagai produk akhir dari gen. RNA ini juga dapat berfungsi seperti protein seperti sebagai katalis berbagai reaksi enzimatik maupun komponen struktural dari berbagai molekul di sel.
Contohnya adalah enzim telomerase, yang merupakan enzim yang tersusun dari RNA. Namun, banyak dari RNA ini masih belum diketahui fungsinya di dalam sel. Beberapa jenis RNA yang telah diketahui adalah sebagai berikut:
| Jenis RNA | Fungsi |
|---|---|
| mRNA | Messenger RNA, kode untuk sintesis protein |
| rRNA | Ribosomal RNA, membentuk struktur dasar dari ribosom dan mengkatalisasi sintesis protein |
| tRNA | Transfer RNA, berfungsis entral dalam sintesis protein sebagai adaptor antara mRNA dengan asam amino |
| snRNA | Small nuclear RNA, berfungsi dalam berbagai proses di inti sel, termasuk membelah pre-mRNA |
| snoRNA | Small nucleolar RNA, digunakan dalam proses dan secara kimiawi memodifikasi rRNA |
| scaRNA | Small cajal RNA, berperan dalam modifikasi snoRNA dan snRNA |
| miRNA | Micro RNA, meregulasi ekspresi gen khususnya dengan memblok translasi dari mRNA tertentu |
| siRNA | Small interfering RNA, mematikan ekspresi gen dengan mengarahkan degradasi dari mRNA tertentu dan membentuk struktur kompak dari kromatin |
| RNA nonkoding lainnya | Berfungsi dalam berbagai macam proses di sel termasuk sintesis telomer, inaktivaso kromosom X, dan transpor protein ke retikulum endoplasma. |
Satu Unit Transkripsi: Apakah Selalu Satu Gen Satu Protein/RNA?
Setiap unit segmen DNA yang ditranskripsikan disebut sebagai satu unit transkripsi. Pada eukariota, satu unit transkripsi umumnya membawa informasi hanya satu gen sehingga mengkode hanya satu molekul RNA atau satu molekul protein saja (atau satu grup protein yang berkaitan jika transkrip RNA dibelah lebih dari satu menghasilkan beberapa mRNA).
Namun, pada bakteri, satu set gen yang berdekatan satu sama lain sering kali ditranskripkan sebagai satu unit. Hasilnya adalah molekul mRNA yang membawa informasi beberapa protein yang berbeda. Jadi, dalam kondisi ini, kondisi satu gen satu protein atau satu RNA tidak berlaku.
Sekuens atau Tahapan Proses Transkripsi DNA
Sekuens atau tahapan transkripsi DNA terdiri dari tiga tahapan utama yaitu:
- Inisiasi, yaitu tahapan awal transkripsi DNA. Proses ini terjadi saat RNA poliemrase berikatan ke bagian promoter dari gen. Kejadian ini merupakan sinyal bagi DNA untuk dibuka sehingga enzim bisa membaca basa dan membuat salinan gen berupa RNA. dari template DNA.
- Elongasi, yaitu proses pemanjangan RNA hasil transkripsi. Satu persatu nukleotida ditambahkan dan RNA bertambah panjang. Nanti akan dijelaskan bahwa pada tahap awal inisiasi, proses pemanjangan RNA akan berjalan lambat. Namun, setelah masuk fase inisiase, pemanjangan ini akan berlangsung lebih cepat.
- Terminasi, yaitu penghentian proses transkripsi. Hal ini terjadi saat RNA polimerase menemui kode codon stop yang menandakan tempat berakhirnya gen.
Pada dasarnya sekuens transkripsi ini sama antara prokariota dengan eukariota. Namun, khusus pada proses inisiasi, pada eukariota berlangsung lebih kompleks. Untuk lebih jelasnya akan dijelaskan masing-masing proses di atas berikut ini:
Sinyal Inisiasi Transkripsi DNA
Agar proses transkripsi berjalan akurat, maka RNA polimerase harus tahu tempat gen dimulai dan tempat gen berakhir. Cara atau proses pengenalan ini berbeda antara bakteri dengan eukariota. Kita akan membahas terlebih dahulu proses RNA polimerase mengenali awal dan akhir dari gen pada bakteri terlebih dahulu.
Tahap inisiasi transkripsi sangat penting pada ekspresi gen karena pada tahap ini sel meregulasi atau mengatur jenis protein apa yang diproduksi dan seberapa banyak produksi yang harus dilakukan.
Seperti disebutkan di atas, untuk proses inisiasi, maka DNA polimerase harus pertama kali menempel ke bagian DNA yang dinamakan promoter. Promoter adalah bagian DNA berupa sekuens atau susunan nukleotida unik yang menandai titik dimulainya transkripsi DNA. Gambar di bawah ini menggambarkan posisi promoter dengan gen yang akan ditranskripsikan.

Tampak di gambar atas bahwa bagian promoter ada di depan gen. Terdapat sekuens khusus yang tersusun dari nukleotida T dan A sehingga regio tersebut dinamakan TATA box. TATA box penting karena di sana adalah tempat menempelnya faktor transkripsi. Selain TAAT box, untuk pengenalan promoter juga terdapat sekuens inisiasi tambahan yang memiliki susunan nukleotida unik yang menjadi petanda bagian promoter dari suatu gen.
Faktor Inisiasi Transkripsi Gen pada Prokariota
Inti enzim dari RNA polimerase bakteri merupakan struktur kompleks multisubunit yang melakukan sintesis RNA dengan DNA sebagai template. Pada proses inisiasi gen, RNA polimerase dibantu oleh protein lain yang dinamakan faktor transkripsi. Pada bakteri, hanya terdapat satu faktor transkripsi yaitu disebut faktor sigma (σ). Faktor σ berkaitan dengan inti enzim dan membantu dalam membaca sinyal di DNA di bagian promoter yang menandakan dimulainya proses transkripsi. Inti enzim RNA polimerase bersama-sama dengan faktor σ dinamai holoenzim RNA polimerase.
Kompleks holoenzim RNA polimerase ini hanya terikat secara lemah dengan DNA bakteri ketika terjadi tumbukan dan kompleks ini akan bergeser dengan cepat sepanjang molekul DNA sampai kemudian berpisah kembali. Tetapi, apabila holoenzim polimerase melalui faktor σ bertemu dengan bagian DNA yang dinamakan promoter, maka akan terjadi ikatan yang kuat antara enzim dengan DNA.
Adapun pada saat mulai terjadinya transkripsi, terjadi proses kompleks berupa perubahan bentuk atau konformasi dari enzim. Kita dapat memandang proses ini sebagai pembukaan dan peletakan DNA di situs aktif dari enzim diikuti dengan penguncian dari enzim sekitar DNA dan RNA. Penguncian ini untuk memastikan struktur tidak berpisah sebelum proses transkripsi selesai. Jika proses transkripsi berhenti di tengah jalan, maka proses inisiasi harus dimulai dari awal kembali yaitu dari bagian promoter.

Inisiasi Transkripsi DNA pada Eukariota
Selain eukariota memiliki RNA polimerase khusus, terdapat perbedaan proses inisiasi transkripsi antara prokariota dengan eukariota. Perbedaan inisiasi ini seperti disebutkan di atas yaitu RNA polimerase II memerlukan banyak tambahan protein yang dinamakan faktor transkripsi umum (general transcription factors) dan inisiasi transkripsi pada eukariota harus berhadapan dengan struktur penyusun nukleosom DNA dan struktur kromatin lainnya yang tidak dijumpai pada kromosom bakteri.
Faktor Transkripsi Umum
Faktor transkripsi umum membantu memposisikan RNA polimerase eukariota secara tepat di promoter, membantu memisahkan dua strand DNA sehingga transkripsi dapat dimulai, dan melepas RNA polimerase dari promoter ke mode elongasi.
Disebut faktor “umum” karena protein dibutuhkan untuk semua promoter yang menggunakan RNA polimerase II. Faktor transkripsi umum ini terdiri dari satu set protein yang saling berinteraksi dan dinamakan TFII (dari kata transcription factor for polymerase II) dan didenotasi secara arbitrer sebagai TFIIB, TFIID, dan seterusnya. Secara umum, TFII ini menjalankan fungsis eperti faktor σ dan bagian dari TFIIF memiliki struktur tiga dimensi yang equivalen dengan faktor σ.
| Nama | Jumlah subunit | Peran pada inisiasi transisi |
|---|---|---|
| TFIID | ||
| – Subunit TBP | 1 | Mengenali TATA box |
| – Subunit TAF | ~11 | Mengenali sekuens lain dekat titik dimulainya transkripsi, mengatur DNA-binding oleh TBP |
| TFIIB | 1 | Mengenali elemen BRE di promoter, secara akurat menempatkan RNA polimerase di tempat mulainya transkripsi |
| TFIIF | 3 | Stabilisasi interaksi RNA polimerase dengan TBP dan RFIIB; membantu menarik TFIIE dan TFIIH |
| TFIIE | 2 | Menarik dan mengatur TFIIH |
| TFIIH | 9 | Membuka heliks DNA di tempat transkripsi, fosforilasi Ser5 dari CTD RNA polimerase, melepas RNA polimerase dari promoter |
Proses penyusunan dimulai dengan ikatan TFIID ke sekuens pendek DNA double heliks yang utamanya tersusun dari nukleotida T dan A sehingga disebut TATA box dan subsunit dari TFIID yang mengenali TATA box dinamai TBP (TATA binding protein).
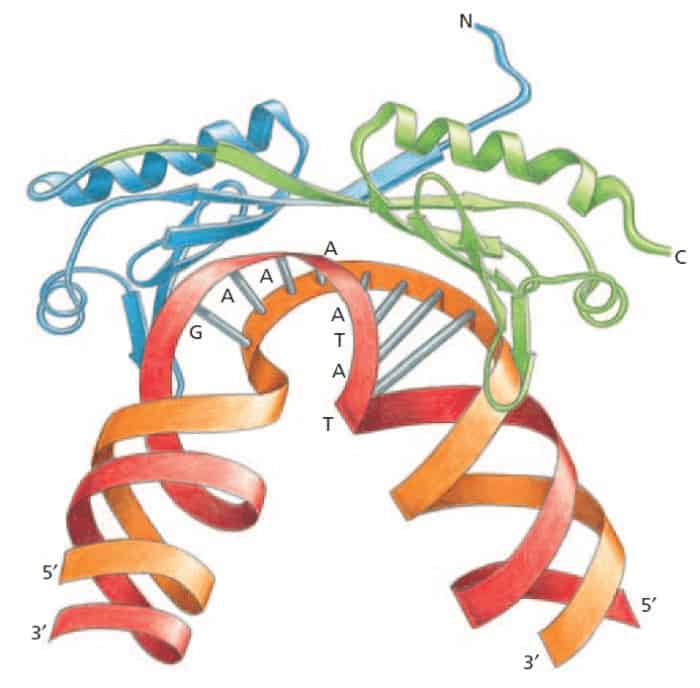
TATA box terletak terutama 25 nukleotida upstream dari tempat transkripsi dimulai. Terdapat sekuens lain yang menjadi sinyal dimulainya transkripsi namun untuk RNA polimerase II, promoter ini adalah promoter yang paling penting.

Ikatan TBP ke TATA box menyebabkan distorsi dari DNA di sekuens ini. Distorsi ini menjadi tanda fisik untuk lokasi promotor yang aktif dan memungkinkan langkah penyusunan protein lain untuk transkripsi dimulai.
Faktor Transkripsi Umum dan RNA Polimerase II Membentuk Transcription Initiation Complex
Faktor transkripsi umum yang disusun bersama-sama dengan RNA polimerase II membentuk kompleks protein besar yang dinamakan transcription initiation complex. Faktor yang paling kompleks adalah TFIIH yang terdiri dari 9 subunit dan dengan besar hampir menyamai RNA polimerase II. TFIIH ini mengandung DNA helikase di satu dari subunitnya dan bekerja dengan menggunakan ATP untuk membongkar struktur double heliks DNA.
Setelah transcription initiation complex terbentuk di promoter DNA, RNA polimerase II harus mendapat akses ke template strand di titik dimulainya transkripsi. TFIIH dengan aktivitasa helikasenya membuat proses ini terjadi.
Selanjutnya RNA polimerase II seperti pada bakteri akan tetap berada di promoter mensintesis RNA pendek sampai kemudian mengalami serangkaian perubahan konformasi yang menyebabkan enzim bergerak dan masuk ke fase elongasi.
Satu langkah kunci pada transisi ini adalah penambahan gugus fosfat di ekor dari RNA polimerase (dikenal sebagai CTD atau C-terminal domain). Pada manusia, CTD terdiri dari 52 tandem repeat dari tujuh sekuens asam amino yang memanjang dari struktur inti RNA polimerase.

Saat inisiasi transkripsi, serine yang berada di posisi kelima dari repeat sequence (Ser5) akan difosforilasi oleh TFIIH. Polimerase kemudian akan terpisah dari cluster general transcription factors. Pada proses ini terjadi perubahan konformasi sehingga enzim akan secara erat berinteraksi dengan DNA dan mendapat protein baru lainnya yang memungkinkan proses transkripsi berlangsung lama dan dapat beberapa jam terus-menerus tanpa lepas dari DNA.
Saat RNA polimerase II memulai elongasi dari transkrip RNA, kebanyakan general transcription factors akan melepaskan diri dari DNA sehingga dapat digunakan kembali dalam proses inisiasi transkripsi berikutnya. Selain itu, fosforilasi ekor dari RNA polimerase juga memungkinkan protein lain yang berperan dalam pemrosesan RNA turut bergabung ke RNA polimerase sehingga menyebabkan RNA yang baru dibentuk untuk diproses dan dimodifikasi.
Polimerase II Membutuhkan Aktivator, Mediator, dan Chromatin-Modifying Proteins
Seperti disebutkan di atas bahwa DNA di sel eukariota dikemas dalam bentuk nukleosom dan dikemas lagi menjadi bentuk yang lebih tinggi yaitu struktur kromatin. Akibatnya, insiasi transkripsi DNA di sel eukariota lebih kompleks dan memerlukan lebih banyak bantuan protein lain.
Pertama, protein regulator gen yang disebut aktivator transkripsional harus mengikat sekuens DNA spesifik dan membantu untuk menarik RNA polimerase ke titik dimulainya transkripsi.
Kedua, secara in vivo, proses transkripsi dalam sel eukariota memerlukan mediator. Mediator memungkinkan beberapa aktivator dapat berkomunikasi baik dengan RNA polimerase II maupun dengan faktor transkripsi umum.
Ketiga, terakhir, proses transkripsi di sel eukariota memerlukan rekruitmen lokal dari chromatin-modifying enzyme baik chromatin remodeling complex maupun histone-modifying enzyme. Keduanya memungkinkan akses yang lebih besar ke DNA yang berada di dalam kromatin sehingga memfasilitasi perakitan kompleks mesin penginisiasi transkripsi DNA.

Banyak protein (lebih dari 100 subunit) harus dirakit di titik tempat mulainya proses transkripsi untuk memulai atau menginisiasi transkripsi di sel eukariota. Urutan perakitan tidak perlu bertahap dan bahkan berbeda dari satu gen ke gen yang lain. Begitu juga proses pengaturan kompleks ini bahkan memerlukan proses seperti proteolisis di tempat untuk melepaskan sub unit tertentu dari kompleks enzim RNA polimerase. Dengan demikian, cara sel meregulasi ekspresi gen sangat banyak dan akan kita bahas di kesempatan khusus di artikel yang lain.
Elongasi Transkripsi DNA Menyebabkan Terbentuknya Superhelical Tension pada DNA
Ketika sudah terinisiasi, kerja RNA polimerase tidak berlangsung dengan halus atau lancar sepanjang DNA. Kadang tersendat, berhenti sementara pada beberapa sekuens dan berjalan cepat di tempat yang lain. RNA polimerase yang sedang dalam fase elongasi berkaitan dengan faktor elongasi yaitu protein yang mengurangi kemungkinan RNA polimerase akan terlepas dari DNA sebelum trenskripsi mencapai titik terminasi.
Faktor elongasi ini berikatan dengan RNA polimerase segera setelah proses inisiasi selesai dan membantu polimerase bergerak sepanjang sekuens template DNA dan struktur kromatin pada eukariota. Dengan memakai ATP, chromatin remodeling complex akan membuka struktur kromatin.
Kompleks ini dapat bergerak bersama dengan polimerase namun dapat pula muncul pada RNA polimerase yang macet dan setelah bergerak kembali akan lepas. Adapun mengenai detil mekanisme ini masih belum terungkap secara lengkap namun diketahui protein tersebut dapat melepas secara sementara dimer H2A-H2B dari inti nukleosom, mengganti mereka saat pomerase bergerak sepanjang nukleosom.
Terdapat hambatan lain dalam proses elongasi yaitu adanya DNA supercoiling. Hal ini terjadi ketika gaya torsi terbentuk saat proses transkripsi DNA berlangsung. Perhatikan gambar di bawah menggambarkan bagaimana DNA supercoiling ini terbentuk:
Untuk mengurangi atau mengatasi supercoiling ini, maka terdapat mekanisme berupa enzim topoisomerase pada eukariota dan bakteri. Namun pada terdapat topoisomerase khusus yang dinamakan DNA gyrase.

DNA gyrase menggunakan ATP secara aktif menciptakan supercoiling di belakang RNA polimerase. Supercoiling yang diciptakan berkebalikan (negative supercoiling) dengan supercoiling yang disebabkan RNA polimerase sehingga efeknya saling menghilangkan.
Negative supercoiling ini makah dapat memfasilitasi proses pembukaan heliks dari DNA sehingga membantu proses transkripsi DNA pada bakteri. Perlu diingat bahwa DNA gyrase tidak ada pada eukariota.
Sinyal Terminasi Transkripsi DNA
Proses termiansi ditandai saat RNA polimerase mengenali sinyal terminasi. Pada kebanyakan dari gen bakteri, sinyal terminasi terdiri dari susunan berulang pasangan nukleotida A-T yang didahului oleh sekuens DNA simetris sepanjang dua kali lipat, dimana jika ditranskripsikan, bagian ini akan membentuk RNA berbentuk jepit rambut atau ‘hairpin‘.
Saat RNA polimerase sampai ke bagian ini, bentuk jepit rambut dari RNA membantu RNA yang terbentuk ditarik dari situs aktif enzim. Hibrid DNA-RNA pada situ aktif yang ditahan bersama oleh pasangan terminator berupa pasangan ikatan U-A tidak terlalu kuat untuk menahan RNA tetap berada di tempatnya sehingga akan menyebabkan RNA terlepas dan akhirnya proses transkripsi berakhir.

Heterogenitas Sekuens Nukleotida Petanda Inisiasi dan Terminasi Transkripsi DNA
Terdapat variasi pola sekuen nukleotida yang menjadi petanda inisiasi dan terminasi proses transkripsi DNA. Di bawah ini adalah contoh variablitias sekuens inisiasi dari bakeri E. coli:

Modifikasi dan Pemrosesan RNA Pasca Transkripsi DNA
Setelah terbentuk RNA, maka pada gen yang mengkodekan protein akan dilanjutkan ke proses translasi RNA. RNA yang ditranslasikan berupa mRNA. Pada prokariota, proses ini berlangsung cukup sederhana. Dikarenakan tidak adanya struktur inti sel, mRNA yang dihasilkan dapat langsung ditranslasikan.
Namun pada euakriota terdapat proses modifikasi terlebih dahulu RNA sebelum ditranslasikan. Selain itu, proses translasi mRNA pada eukariota berlangsung di sitoplasma sehingga mRNA harus diekspor terlebih dahulu keluar dari inti sel. Perhatikan gambar di bawah ini:

Jadi pada gambar tampak bahwa transkripsi DNA pada eukariota hanya merupakan langkah pertama dari langkah lanjutan untuk memproduksi mRNA. Langkah lain berupa modifikasi secara kovalen kedua ujung RNA dan pembuangan sekuens intron.
Mekanisme transkripsi dan pemrosesan ini dikemas dengan sangat baik oleh sel. Salah satu kunci adalah fosforilasi CTD. Selain membantu proses pelepasan RNA dari template DNA, CTD juga menjadi kerangkan bagi protein yang terlibat dalam pemrosesan RNA untuk bergabung ke kompleks repliklasi dan melakukan modifikasi RNA on the spot.
Di bagian kali ini kita akan membahas modifikasi RNA pada eukariota secara lebih detil sampai pada proses terbentuknya mRNA yang matur.
RNA Capping Merupakan Modifikasi Pertama dari Pre-mRNA Eukariota

Segera setelah diproduksi kurang lebih 25 nukelotida, maka ujung 5′ dari RNA yang baru akan ditambahkan cap atau pelindung. Cap ini berupa molekul guanine yang dimodifikasi. Tiga enzim bekerja secara berurutan dalam proses ini:
- Enzim fosfatase membuang satu gugus fosfat di ujung 5′ dari RNA yang baru terbentuk
- Enzim guanil transferase menambah GMP dengan ikatan terbalik (5′ ke 5′ dari pada 5′ ke 3′)
- Enzim metil transferase menambah gugus metil ke guanosine.

Perlu diingat bahwa ketiga enzim ini terikat ekor RNA polimerase (CTD) yang terfosforilasi di posisi Ser5. Dengan demikian modifikasi ini segera dilakukan setelah RNA yang diproduksi ujungnya keluar dari situs katalitik RNA polimerase.

Modifikasi ini menyebabkan mRNA dapat lebih mudah dikenali dan dibedakan dengan tipe RNA lainnya oleh sel. RNA polimerase I dan III tidak memiliki CTD sehingga RNA yang diproduksi tidak memiliki cap.
Di inti sel, sekelompok kompleks protein akan mengikat cap dan disebut CPC (cap-binding complex). CPC ini membantu RNA yang dihasilkan diproses dan ditransportasikan dengan benar. Selain itu, gugus 5′-methyl pada cap juga membantu proses translasi mRNA di sitoplasma.
RNA Splicing Membuang Sekuens Intron dari Pre-mRNA
Seperti disebutkan di atas bahwa pada segmen gen pada eukariota terdapat sekuens non koding yang disebut intron. Intron ditemukan pada tahun 1977 dan cukup mengejutkan para ahli karena waktu itu hanya familiar dengan gen bakteri yang tidak memiliki intron. Adapun bagian gen yang mengkodekan protein disebut ekson dan kadang pada satu gen bagian ekson hanya mencakup sebagian kecil dari keseluruhan gen.
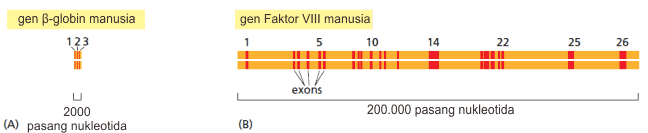
Pada saat transkripsi, baik intron maupun ekson disalin menjadi RNA yang baru. Namun, kemudian bagian intron akan digunti keluar melalui proses RNA splicing. Supaya tidak membingungkan, kita memakai istilah precursor-mRNA (pre-mRNA) untuk RNA yang baru diproduksi sedangkan mRNA untuk RNA yang telah sudah dimodifikasi di ujung 5′ dan 3′ dan juga telah mengalami RNA splicing.
Tiap peristiwa RNA splicing menggambarkan proses pembuangan satu intron. Proses ini terdiri dari dua reaksi fosforil-transfer secara berurutan yang disebut transesterifikasi. Transesterifikasi ini menghilangkan intron dan kemudian membentuk “lariat”.

Struktur yang terlibat dalam proses RNA splicing ini terdiri dari komponen molekul kompleks yang terdiri dari lima potong RNA dan sekitar 200 protein. Untuk setiap proses splicing, menghidrolisis banyak molekul ATP. Kompleksitas ini menjamin bahwa proses RNA splicing berjalan secara akurat.
Kenapa Terdapat Intron pada Eukariota?
Dengan adanya banyak bagian intron yang dibuang tampaknya mesin DNA kurang efisien. Salah satu penjelasan kenapa banyak segmen DNA berupa intron, para ahli melihatnya dari sisi proses evolusi.
Susunan intron-ekson dilihat bisa memfasilitasi kemunculan protein baru yang berguna pada proses evolusi seiring waktu. Keberadaan banyak intron memungkinkan rekombinasi genetik membentuk ekson dari gen-gen yang berbeda dan terbentuklah gen dari protein baru yang memungkinkan proses evolusi. Apabila kita lihat, gen yang ada saat ini tampak seperti susunan berbagai jenis domain protein yang lebih sederhana sehingga mendukung ide bahwa adanya intron memfasilitasi pembentukan protein baru.
Dalam proses biologi selain evolusi, keberada intron juga memberikan manfaat. Transkrip dari banyak gen eukariota (75% pada gen manusia) dipotong lebih dari satu cara sehingga memungkinkan satu gen untuk menghasilkan satu set protein-protein yang berbeda. Dengan adanya splicing ini memungkinkan eukariota untuk meningkatkan potensi koding dari gen yang dimiliki.

Sinyal Nukleotida Penanda Tempat RNA Slicing
Agar proses RNA splicing dapat terjadi secara akurat, maka protein yang melakukan splicing ini harus mengenali tempat 5′ dari proses splicing, tempat 3′, dan titik percabangan dari sekuens intron. Tiap titik ini memiliki pola sekuens nukleotida konsensus yang menjadi petanda tempat terjadinya RNA splicing. Sekuens petanda ini umumnya pendek dan juga terdapat variabilitas.

RNA Splicing Dilakukan oleh Spliceosome
Tidak seperti proses tahapan mRNA sebelumnya, RNA splicing dilakukan oleh molekul RNA. Molekul RNA khusus melakukan pengenalan sekuens nukleotida tempat dilakukannya splicing dan juga melakukan reaksi kimia yang memotong pre-mRNA tersebut.
Molekul RNA ini umumnya relatif pendek (kurang dari 200 nukleotida) dan terdapat lima molekul yang dinamai U1, U2, U4, U5, dan U6. Molekul RNA ini masuk dalam kelompok tersendiri yaitu snRNA (small nuclear RNA). Setiap snRNA ini dilengkapi dengan tuhjuh protein membentuk snRNP (small nuclear ribonucleoprotein). Tiap snRNP ini membentuk inti dari spliceosome, susunan besar RNA dan protein yang melakukan splicing pre-mRNA.
Spiceosome ini kompels dan dinamik. Pada studi in vitro , beberapa komponen akan terususn di sekitar pre-mRNA dan kemudian komponen lain akan saling berdatangan untuk melakukan tugasnya. Namun, para ahli berpendapat bahwa dalam sel, nucleosome ini sudah terakit dan siap melakukan kerjanya di dalam inti sel.
Pada saat terjadi proses splicing, bentuk spliceosome akan berubah dan terjadi pergeseran banyak komponen. Sebagai contoh U1 akan diganti oleh U6 ditempat 5′ terjadinya pemotongan RNA. Pergantian susunan ini terjadi beberapa kali dan hal ini memungkinkan terjadinya check dan recheck untuk meningkatkan akurasi RNA splicing.

Spliceosome Mengunakan Hidrolisis ATP Menghasilkan Serangkaian Kompleks Susunan RNA-RNA
Walaupun ATP tidak diperlukan dalam reaksi kimia pemotongan pre-mRNA, namun ATP diperlukan dalam menyusun spliceosome. Protein-protein tambahan yang menyusun spliceosome menggunakan ATP untuk memecah interaksi RNA-RNA sehingga memungkinkan perubahan formasi yang baru.
Salah satu fungsi penting dalam penyusunan dan perubahan interaksi RNA-RNA ini adalah untuk membentuk situs aktif pembelahan. Dibentuknya situs aktif setelah komponen lain terbentuk merupakan strategi untuk mencegah terjadinya pemotongan pre-mRNA yang serampangan.

Saat proses pemotongan selesai, snRNP akan tetap terikat pada lariat. Pembongkaran snRNP dari lariat memerlukan perubahan formasi RNA-RNA lanjutan yang membutuhkan ATP untuk mengembalikan susunan snRNP seperti semula. Di tempat titik pemotongan, spliceosome juga mengarahkan kelompok protein lain untuk mengikat mRNA di tempat pemotongan. Protein ini dinamakan exon junction complex (EJC) yang menandai proses splicing yang telah selesai. EJC ini akan berperan dalam proses perjalanan mRNA selanjutnya.
Sintesis dan Sifat Lainnya dari Pre-mRNA Menjelaskan Pilihan Tempat Terjadinya RNA Splicing
Sekuens intron sangat bervariasi dalam hal ukuran dengan beberapa mencapai 100.000 nukleotida. Apabila pemilihan tempat splicing hanya mengandalkan kerja snRNP maka diperkirakan akan terjadi banyak sekali kesalahan pemotongan.

selection
Ketepatan spliceosome dalam mengerjakan tugasnya dibantu dua tambahan strategi. Pertama adalah sebagai konsekuensi bahwa proses splicing terjadi bersamaan saat RNA disintesis oleh RNA polimerase II. Hal ini membuat mekanisme sel dapat mengontrol dan membaca bagian intron dan ekson secara lengkap.
Strategi kedua adalah apa yang disebut dengan definisi ekson. Besarnya ekson cenderung lebih seragam dibandingkan intron dengan rata-rata 150 pasang basa. Dengan proses ini, mekanisme sel akan mencari sekuens ekson yang realtif serupa.

Ketika proses sintesis RNA berjalan, satu grup komponen tambahan (terutama protein SR dinamakan demikian karena mengandung domain yang kaya serine dan arginine) akan tersusun di sekuens ekson dan membantu menandai setiap ujung 3′ dan 5′ tempat pemotongan. Protein ini kemudian akan merekruit U1 snRNA yang menandai batas donwstream dari ekson dan U2AF yang menandai batas usptream dari ekson.

Detil lebih jauh mengenai bagaimana mekanisme protein SR membedakan intron dengan ekson masih belum dipahami dengan jelas. Namun diketahui bahwa beberapa protein SR mengikat secara khusus ke sekuens RNA spesifik di ekson yang dikenal sebagai splicing enhancers.
Set Kedua dari snRNP Memotong Sebagian Kecil Sekuens Intron pada Hewan dan Tanaman
Eukariota sederhana seperti ragi hanya memiliki satu set snRNP yang mengerjalan seluruh reaksi RNA splicing. Pada organisme yang lebih kompleks seperti lalat, mamalia, dan tanaman memiliki set kedua dari snRNP yang memotong langsung sebagian kecil sekuens intron. Bentuk minor dari spliceosome ini mengenal set sekuens RNA yang berbeda di titik 3′ dan 5′ pemotongan dan percabangan. Set snRNP ini disebut U12-type spliceosome.
Walaupun mengenai sekuens nukleotida yang berbeda, spliceosome ini juga membuat jenis interaksi RNA-RNA yang sama dengan tipe snRNP mayor lainnya. Namun, U12-type spliceosome memiliki perbedaan komponen spliceosome yang berjalan bersama-sama dengan RNA polimerase II. Selain itu, pada snRNP tipe ini, waktu pemotongan dapat terjadi lebih lambat. Hal ini memungkinkan sel untuk meregulasi ekspresi gen yang memerlukan pemotongan pre-mRNA dengan jenis spliceosome ini,
Beberapa organime eukariota juga memiliki variasi splicing dinamakan trans-splicing. Organisme ini termasuk jenis trypanosoma dan cacing nematoda. Pada trans-splicing, ekson dari dua transkrip RNA yang berbeda dipotong bersama-sama dan membentuk satu molekul mRNA yang matur.
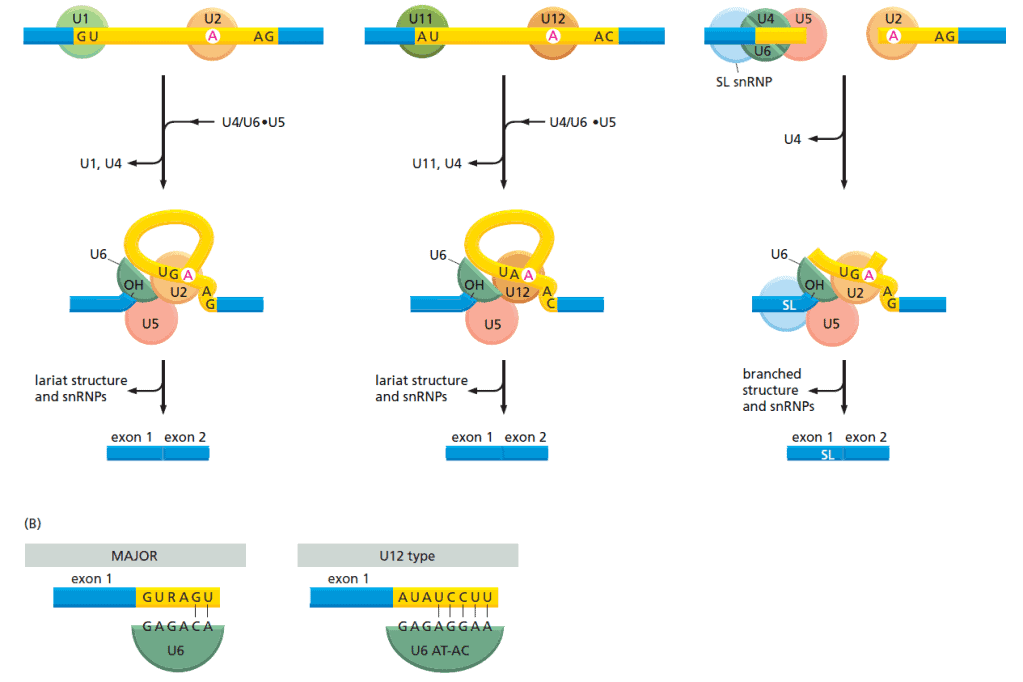
Trypanosoma memproduksi semua mRNA dengan cara ini sedangkan pada nematoda hanya 1% saja diproduksi dengan trans-splicing. Pada kedua organisme ini, satu ekson dipotong dan digabungkan dengan ujung 5′ dari berbagai gen yang lain. Hal ini menyebabkan banyak mRNA yang memiliki ujung 5′ yang sama namun ujung 3′ yang berbeda. Trans-splicing ini dikerjakan oleh snRNP khusus yang dinamakan SL RNP.
Kita tidak tahu kenapa ada organisme yang menggunakan proses trans-splicing. Namun, diperkirakan bahwa dengan adanya kesamaan ujung 5′, akan membantu proses translasi dari mRNA. Hal ini diperkuat dengan mRNA hasil trans-splicing pada nematoda yang ditranslasikan dengan tingkat efisiensi yang tinggi.
Plastisitas pada RNA Splicing
Dari penjelasan di atas bahwa diperlukan tiga petanda untuk menentukan letak pemotongan intron yaitu sinyal di RNA (ujung 3′, ujung 5′, dan tempat percabangan), penyusunan kotranskripsional dari spliceosome, dan mekanisme pengenalan ekson. Kita tidak mengetahui seberapa tepat akurasi sistem pemotongan intron karena terdapat mekanisme penghancuran mRNA yang salah produksi. Namun, kita mengetahui bahwa dibandingkan proses lain, mekanisme splicing ternyata sangat fleksibel.
Sebagai contoh adalah mutasi pada sekuens nukleotida yang penting untuk proses splicing namun tidak menyebabkan terhentinya proses splicing secara total. Hal ini menyebabkan terjadinya pola baru proses splicing. Lihat contohnya di bawah ini:

Paling banyak ditemui adalah satu ekson diloncat. Pada kasus lain, mutasi menyebabkan cryptic splice junction. Ternyata mekanisme splicing berevolusi sehingga memilih pola terbaik untuk melakukan pemotongan. Apabila mutasi menyebabkan titik pemotongan paling optimal rusak, maka mekanisme pemotongan akan memilih titik lain yang paling optimal.
Fleksibilitas ini memberi petanda bahwa proses evolusi dimungkinkan dengan adanya mutasi. Selain itu mekanisme ini menyebabkan sel mampu meregulasi pola RNA splicing. Kita akan membicarakan mekanisme pengaturan ini di artikel yang membahas mengenai mekanisme regulasi ekspresi gen.
Spliceosome Berevolusi dari Mekanisme Self-Splicing
Mekanisme splicing dari pre-mRNA diperkirakan berkembang dan berevolusi dari proses yang lebih sederhana. Proses yang sederhana ini berupa self-splicing, artinya intron dapat melakukan pemotongan oleh dirinya sendiri. Hal ini terobservasi pada gen inti rRNA dari siliata Tetrahymena, beberapa gen bakteriofag T4, dan beberapa gen mitokondria serta kloroplas.
Sekuens intron yang memiliki aktivitas self-splicing dapat diidentifikasi di cawan petri dengan melakukan inkubasi molekul RNA murni dan melihat apakah terjadi reaksi RNA splicing. Dengan cara ini, ditemukan dua kelas utama self-splicing intron:
- Sekuens intron grup I memulai reaksi dengan mengikat nukleotida G pada sekuens intron. Nukleotida ini menjadi grup aktif yang menyerang dan memecah ikatan fosfodiester pertama saat proses splicing (titik posisi 5′).
- Sekuens intron grup II, dimana terdapat residu reaktif A sebagai gugus aktif yang memecah ikatan fosfodiester.
Baik grup I maupun grup kemudian menjalani reaksi pemotongan yang kedua dengan proses yang serupa. Perhatikan gambar di bawah ini:
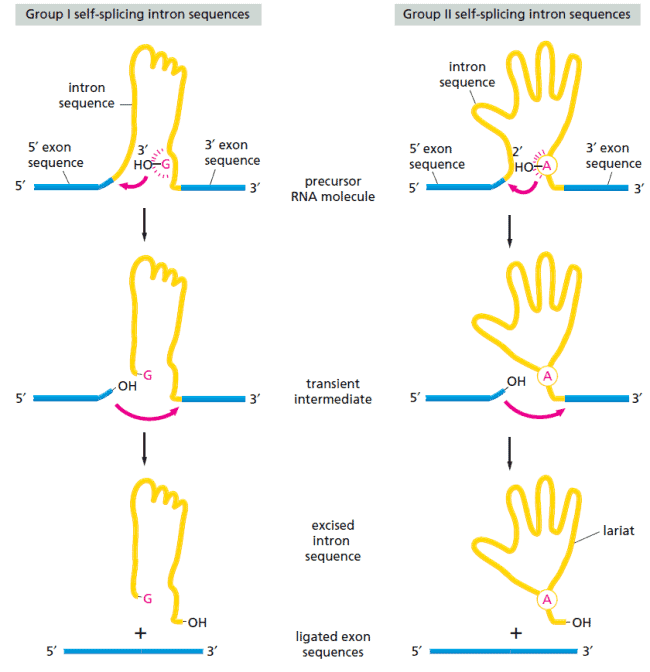
Di kedua grup ini, sekuens nukleotida intron sangat penting karena menentukan bentuk lipatan spesifik dari intron. Tanpa lipatan yang benar, maka reaksi splicing tidak akan terjadi.
Pembentukan Ujung 3′ dari mRNA Eukariota
Perubahan ketiga dan merupakan modifikasi terakhir pre-mRNA pada eukariota adalah perubahan di ujung 3′ dari RNA. Perubahan ini berupa penambahan gugus adenine di ujung 3′.
Seperti dijelaskan sebelumnya bahwa terdapat sinyak khusus yang menjadi petanda akhir gen yang menjadi panduan bagi RNA polimerase untuk menghentikan proses transkripsi DNA. Area ini dikenali oleh protein khusus yaitu dua protein multisubunit yang bernama CstF (cleavage stimulating factor) dan CPSF (cleavage and polyadenylation specific factor). Kedua grup protein ini berjalan seiring dengan RNA polimerase II dan ditransferkan ke ujung 3′ dari sekuens RNA.

Seperti diperkirakan, pengikatan kedua faktor ini ke RNA juga terjadi dengan proses mengenali sekuens spesifik. Saat kedua faktor ini mengenali dan menempel ke RNA, maka protein lain ikut bergabung untuk memproses ujing 3′ dari mRNA. Proses ini adalah sebagai berikut:
- Sebagian ujung RNA dibuang
- Muncul enzim poli-A polimerase (PAP) yang menambahkan gugus adenil sekitar 200 nukleotida adenine di ujung 3′. Enzim PAP ini tidak seperti RNA polimerase karena tidak memerlukan template DNA. Oleh sebab itu, ujung poli-A ini tidak terdapat atau dikodekan di DNA.
Setelah ekor poli-A ini terbentuk, maka datang protein yang bernama poly-A binding protein. Beberapa protein ini akan tetap berada di ekor poli-A dan berjalan bersama mRNA membantu mengarahkan proses sintesis protein di ribosom.
Setelah ujung 3′ dari RNA dilepas, RNA polimerase II akan tetap melakukan proses transkripsi DNA dalam beberapa kasus sampai beberapa ratus nukleotida. Namun, RNA baruini tidak memiliki cap di ujung 5′ dan dengan segera akan dihancurkan oleh eksonuklease 5′ ke 3′. Proses penghancuran ini ternyata memperantarai proses RNA polimerase dan terlepas dari DNA.

Produk Akhir mRNA Diekspor dari Nukleus ke Sitoplasma
Saat mRNA yang matur selesai diproses, maka terdapat perbedaan molekul antara pre-mRNA dengan mRNA yang matur. Perbedaan itu diantaranya adalah protein yang berasosiasi dengan kedua jenis RNA tersebut. Dengan mengenali jenis protein ini, maka sel dapat membedakan antara pre-mRNA dengan mRNA. Hanya mRNA yang matur akan dikirim ke sitoplasma sedangkan sisa RNA, mRNA yang rusak, atau pre-mRNA yang masih diproses akan tetap berada di inti sel. Sebagian dari RNA ini kemudian akan dihancurkan oleh eksosome inti yang berupa protein kompleks besar yang di dalamnya terdapat banyak eksonuklease RNA 3′ ke 5′.
Beberapa jenis molekul yang bernama hnRNP (heterogeneous nuclear ribonuclear protein) berasosiasi dengan pre-mRNA, membuka struktur heliks hairpin dari RNA agar memudahkan pembacaan dan pemrosesan RNA lebih mudah. Pada manusia terdapat sekitar 30 jenis hnRNP. Selain itu protein ini juga membantu membedakan antara mRNA dengan debris sisa transkripsi.
Transkrip mRNA matur yang selesai diproses akan diantar ke nuclear pore complex (NPC). NPC adalah kanal di membran inti yang secara langsung menghubungkan nukleoplasma dengan sitosol. Molekul kecil (< 50.000 dalton) dan berdifusi bebas melalui kanal ini. Namun, makromolekul besar termasuk mRNA yang berasosiasi dengan protein terlalu besar untuk melewati NPC dengan difusi. Sel menggunakan energi untuk secara aktif mentranspor makromolekul tersebut.

Molekul mRNA yang akan diekspor akan disusun dengan protein-protein yaitu hnRNP, protein SR, dan komponen dari spliceosome. Kompleks protein-RNA ini akan mengalami transisi perubahan bentuk menjadi seperti setengah bola melengkung. Bentuk ini kemudian bergerak melalui nukleoplasma dan masuk ke NPC dengan ujung cap 5′ terlebih dahulu dan secara bertahap dilewatkan melalui NPC ke sitoplasma.

Transkripsi DNA dan Pemrosesan RNA Non-Koding di Nukleus
RNA mewakili beberapa persen dari berat kering sel mamalia. Diantaranya hanya 3 – 5% saja yang berupa mRNA yang hanya berupa sebagian kecil dibandingkan dengan sekuens intron yang dibuang. Adapun jenis RNA yang paling banyak terdapat di sel adalah RNA ribosomal (rRNA) yang dapat mencapai 80% dari total RNA di sel yang aktif membelah.
Di sel eukariota, rRNA ini ditranskripsikan oleh tipe polimerase khusus yaitu RNA polimerase I. Struktur kedua polimerase ini sama namun RNA polimerase I tidak memiliki CTD sehingga RNA yang dihasilkan tidak terdapat cap maupun ekor poli-A.
Berbeda dari protein dimana satu gen dapat diamplifiaksi produksinya, rRNA merupakan produk akhir sehingga memerlukan banyak salinan gen untuk memproduksi rRNA dalam jumlah besar. Sel mamalia yang aktif tumbuh memerlukan sekita 10 juta salinan rRNA untuk setiap kali membelah.

Untuk memenuhi kebutuhan ini, sel memang memiliki banyak salinan gen rRNA. Bahkan E. coli mempunyai 7 salinan gen dari rRNA. Adapun manusia memiliki sekitar 200 gen rRNA untuk tiap sel haploid, tersebar dalam satu cluster di lima kromosom yang berbeda. Adapun kodok Xenopus memiliki sekitar 600 salinan gen rRNA setiap genom haploid di satu cluster pada satu kromosom.
Terdapat empat jenis rRNA di sel eukariota dimana setiap tipe berada di tiap ribosom. Tiga dari empat rRNA (18S, 5.8S, dan 28S) dibuat dengan modifikasi kimia dan dari pembelahan saru prekursor rRNA. Adapun rRNA yang keempat (5S) disintesis di cluster gen yang berbeda oleh RNA polimerase III dan tidak memerukan modifikasi.
Fungsi snoRNA dalam Pematangan rRNA
Modifikasi ekstensif terjadi pada prekursor rRNA sepanjang 13.000 nukleotida sebelum dibelah dan disusun menjadi ribosom. Modifikasi ini termasuk 100 metilasi dari posisi 2′-OH gula nukleotida dan 100 isomerasi dari nukleotida uridine menjadi pesudouridine. Fungsi modifikasi ini secara detil masih belum diketahui namun mungkin membantu dalam pelipatan maupun perakitan bentuk rRNA final dan beberapa mempengaruhi fungsi dari ribosom.

Setiap modifikasi ini dibuat pada nukleotida yang spesifik sepanjang prekursor rRNA oleh sekitar 150 “RNA pembimbing”. RNA lain memandu proses pembelahan prekursor rRNA menjadi rRNA yang matur. Semua RNA pemandu ini masuk ke dalam kelompok RNA khusus yang inamakan small nucleolar RNA (snoRNA). Dinamakan demikian karena RNA ini bekerja di kompartemen khusus di dalam inti sel yaitu di nukleolus (anak inti sel).

Banyak dari snoRNA dikodekan dari intron dari gen lain terutama gen yang mengkodekan protein ribosomal. Oleh sebab itu, snoRNA dibuat oleh RNA polimerase II dan dihasilkan dari proses splicing intron dari pre-mRNA.
Selain itu diketahui bahwa beberapa snoRNA-like RNA hanya disintesis oleh sel di otak. Diyakini RNA ini merupakan modifikasi langsung dari mRNA dari pada dari rRNA. Namun, fungsinya terutama dalam regulasi ekspresi gen masih belum dipahami dengan jelas.
Nukleus Merupakan Pabrik Ribosom
Inti sel atau nukleus merupakan organel yang paling jelas tampak di sel eukariota di bawah mikroskop cahaya. Kita tahu bahwa inti sel merupakan tempat pemrosesan rRNA dan perakitannya menjadi subunit ribosom. Namun, tidak seperti organel mayor lainnya, nukleus tidak terikat pada sebuah membran namun berupa agregat makromolekul berupa gen rRNA, prekursor rRNA, rRNA matur, enzim pemrosesan rRNA, snoRNP, protein ribosomal, dan ribosom yang dalam proses perakitan. Asosiasi yang dekat dari semua komponen ini memungkinkan ribosom dirakit dan dibongkar secara cepat dan lancar.
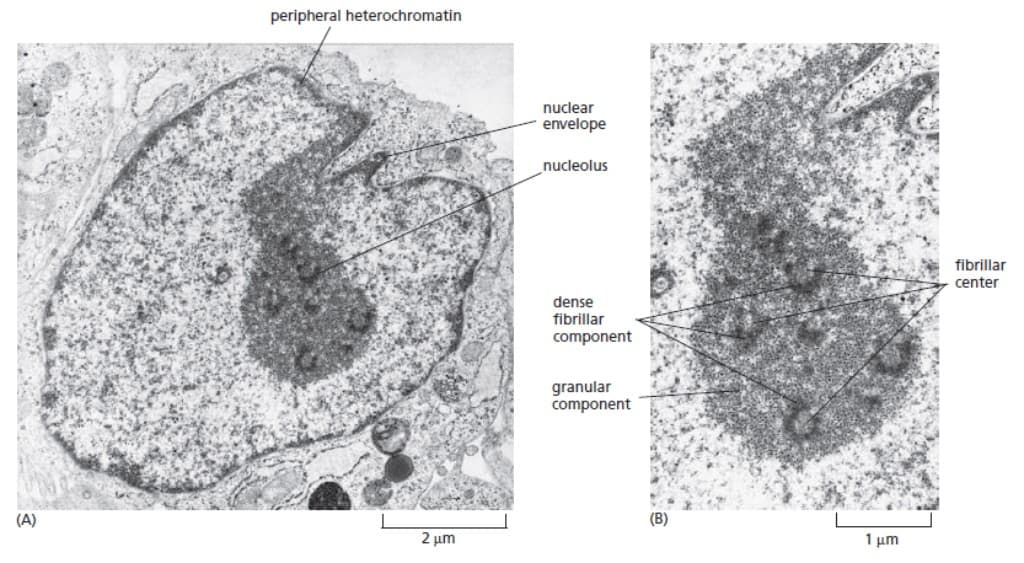
Berbagai jenis tipe RNA memegang peranan penting dalam strktur maupun proses kimiawi di nukleus. Hal ini memberi petunjuk bahwa barang kali nukleus berevolusi dari struktur kuno dari sel dimana reaksi kimia didominasi oleh katalis RNA.
Pada sel diploid manusia, terdapat 10 cluster gen yang mengkodekan rRNA di 5 pasang kromosom yang berbeda. Saat interfase, 10 kromosom ini berkontribusi pada struktur DNA loops pada nukleolus. Pada fase M, saat kromosom berkondensasi, nukleolus menghilang. Di telofase, saat kromosom kembali ke dalam bentuknya yang terdispersi, ujung 10 kromosom ini akan bergabung dan nukleolus akan kembali terbentuk.

Dikarenakan nukleolus menggambarkan banyaknya aktivitas dari pembentukan rRNA, maka dapat diperkirakan bahwa besarnya nukleolus menggambarkan banyaknya ribosom yang diproduksi sel.

Adapun proses perakitan ribosom sendiri merupakan proses yang kompleks. Selain berperan penting dalam pembentukan ribosom, inti sel juga merupakan tempat dimana jenis RNA lain diproduksi dan kompleks RNA-protein lain dirakit. Sebagai contoh, U6 snRNP yang berfungsi dalam RNA splicing. Kompleks protein-RNA lainnya yang penting adalah telomerase dan signal recognition particle. Terakhir, tRNA atau transfer RNA yang membawa asam amino dikodekan oleh cluster gen tRNA di nukleolus.
Nukleus Mengandung Berbagai Struktur Subnuklear
Walaupun nukleolus atau inti sel merupakan struktur yang paling jelas terlihat di inti sel, namun ada beberapa struktur lain di inti sel yang telah dipelajari. Beberapa struktur ini seperti jisim Cajal, GEMS (Gemini of Cajal bodies), dan cluster granula interkromatin (juga dikenal sebagai speckle).
Seperti nukleolus, struktur-struktur tersebut juga tidak terikat struktur membran dan sangat dinamis. Struktur tersebut muncul sebagai hasil kaitan antara protein dengan komponen RNA yang terlibat sintesis, perakitan, dan penyimpanan makromolekul yang terlibat dalam ekspresi gen.

Jisim Cajal dan GEMS mirip satu dengan lainnya dan seringkali berpasangan di nukleus. Masih belum jelas apakah kedua struktur ini merupakan dua struktur yang berlainan. Kedua struktur ini sangat mungkin merupakan lokasi snoRNA dan snRNA mengalami modifikasi kovalen dan menjalani perakitan akhir dengan protein.
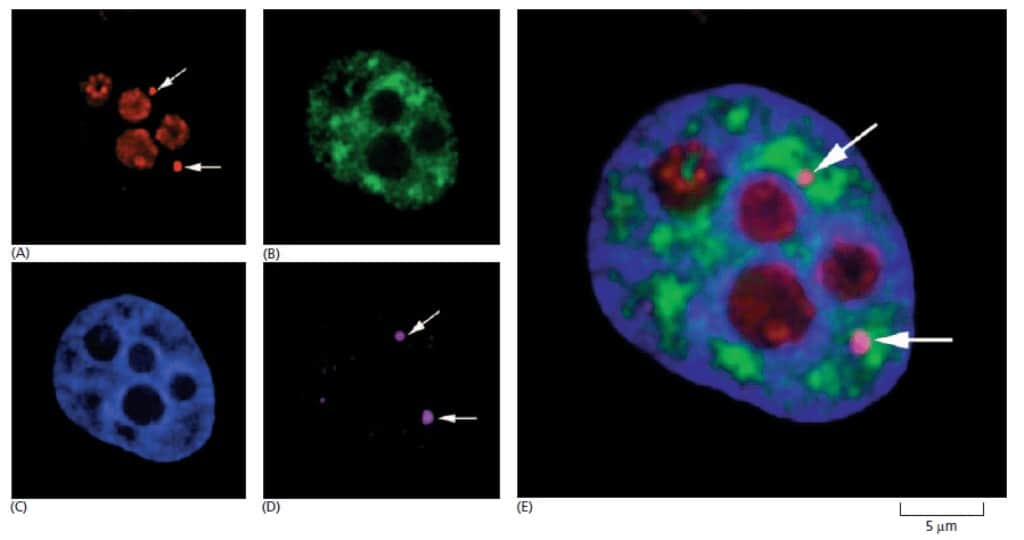
Satu kelompok RNA pembimbing dinamakan small Cajal RNA (scaRNA) memilih lokasi modifikasi ini melalui proses pemasangan basa nukleotida. Jisim Cajal/GEMS juga diperkirakan menjadi tempat dimana snRNP didaur ulang dan susunan RNA-nya direset ulang setelah mengalami perubahan susuna dalam proses RNA splicing.
Penelitian mengenai struktur-struktur tersebut cukup sulit karena penampakan struktur yang berbeda antar organisme dan dapat berubah secara cepat sesuai dengan siklus sel atau dalam respon terhadap perubahan lingkungan.

Pertanyaan-Pertanyaan Seputar Transkripsi DNA (FAQs)
Apa itu proses transkripsi DNA?
Transkripsi DNA adalah proses penyalinan ulang DNA ke molekul lain yaitu RNA.
Proses transkripsi DNA terjadi di mana?
Pada prokariota seperti bakteri, dikarenakan tidak memiliki inti sel maka proses transkripsi DNA terjadi di stoplasma. Sedangkan pada eukariota, transkripsi DNA terjadi di nukleoplasma atau inti sel.
Molekul apa yang dihasilkan dari proses transkripsi DNA?
Transkripsi DNA menghasilkan molekul RNA. RNA ini terdiri dari beberapa jenis. Kebanyakan adalah mRNA yang digunakan lebih lanjut untuk translasi RNA dan sitesis protein. RNA bentuk lain yaitu rRNA, tRNA, snoRNA, dsb.
Apa fungsi proses transkripsi DNA?
Transkripsi DNA berperan dalam proses ekspresi gen. mRNA akan memiliki hasil akhir berupa protein. rRNA dan tRNA berperan dalam membantu proses sintesis proten. Bentuk RNA lain dapat berfungsi sebagai ribozim, pembentuk struktur sel (misalnya ribosom), dan regulasi banyak proses dalam sel.
Enzim apa yang berperan dalam proses transkripsi DNA?
Enzim utama yang berperan dalam transkripsi DNA adalah RNA polimerase.
Apa bedanya transkripsi dengan replikasi DNA?
Proses replikasi DNA adalah penggandaan DNA, jadi DNA disalin menjadi DNA salinannya dan terjadi terutama saat terjadi pembelahan sel. Sedangkan transkripsi adalah penyalinan sebagian dari DNA (biasanya satu gen) menjadi bentuk lain RNA dan berfungsi untuk ekspresi gen.
Kesimpulan
Sebelum dilakukannya sintesis protein, gen yang mengkodekan protein tersebut harus ditranskripsikan terlebih dahulu menjadi mRNA. Terdapat perbedaan yang mencolok antara proses transkripsi DNA pada prokariota dengan eukariota. Pada eukariota, RNA diproduksi oleh tiga tipe RNA polimerase yang berbeda. Selain itu, mRNA yang akan digunakan dalam sintesis protein pada eukariota harus dimodifikasi terlebih dahulu dan diekspor dari inti sel ke sitoplasma sebelum menjalani proses translasi RNA menjadi protein.
Referensi
- Achar YJ, Foiani M. Coordinating Replication with Transcription. In: Advances in Experimental Medicine and Biology. 2017. p. 455–87.
- Aguilera A, Gaillard H. Transcription and Recombination: When RNA Meets DNA. Cold Spring Harb Perspect Biol. 2014;6(8):1–16.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molecular biology of the cell. 5th ed. New York: Garland Science; 2008. 263–328 p.
- Borukhov S, Nudler E. RNA polymerase: the vehicle of transcription. Trends Microbiol. 2008;16(3):126–34.
- Case BC, Hingorani MM. Polymerase ☆. In: Reference Module in Life Sciences. Elsevier; 2017. p. 1–4.
- Dai X, Rothman-Denes LB. DNA structure and transcription. Curr Opin Microbiol. 1999;2(2):126–30.
- Granner DK, Weil PA. RNA synthesis, processing, and modification. In: Murray RK, Granner DK, Mayes PA, Rodwell VW, editors. Harper’s illustrated biochemistry. 26th ed. New York: McGraw-Hill; 2003. p. 341–57.
- Griesenbeck J, Tschochner H, Grohmann D. Structure and Function of RNA Polymerases and the Transcription Machineries. In 2017. p. 225–70.

Seorang dokter, saat ini sedang menjalani pendidikan dokter spesialis penyakit dalam FKUI. Peminat berbagai topik sejarah dan astronomi.
Comments 7
Terima kasih banyak pak, penjelasannya lengkap dan sangat membantu
Terima kasih banyak Pak, sangat bermanfaat sekali untuk saya dan teman2 saya
Terimakasi atas ilmu dan penjelasannya yang sangat lengkap. Sangat membantu saya untuk presentasi materi dikelas nanti.
makasih banyak pak, aslii keren pake banget artikelnya, detail bangett, kalau boleh request pak, transpor elektron dalam respirasi itu bagaimna pak masih sedikit abu-bau buat saya, terimakasih pak saya akan tunggu artikel-artikel bapak selanjuttnya
Author
Artikelnya sudah ada: https://caiherang.com/rantai-transpor-elektron-dan-fosforilasi-oksidatif/. Posisi rantai transpor elektron itu kalau sepemahaman saya mengolah NADH dan FADH2. NADH dan FADH2 datang dari proses siklus Krebs dan glikolisis.
Cuma rantai transpor elektron tidak langsung menghasilkan ATP. Yang dihasilkan adalah energi untuk memompa ion hidrogen dari matriks ke ruang intermembran. Nah, akibatnya ruang konsentrasi ion hidrogen ini tinggi di ruang intermembran. Mitokondria kemudian menggunakan ATP-sintase agar hidrogen yang banyak di ruang intermembran masuk kembali ke dalam matriks.
Nah, sambil lewat melalui ATP-sintase, pergerakan ion hidrogen ini kemudian dipakai untuk produksi ATP. Proses ini dimakan kemiosmosis. Rantai transpor elektron yang diikuti kemiosmosis ini adalah proses yang kita namakan fosforilasi oksidatif.
Terima kasih, dok. Mohon izin untuk dijadikan sumber referensi.
Author
Boleh atau supaya lebih resmi dipilih dari salah satu sumber yg saya cantumkan di bagian referensi.